Download
1 / 12
140 likes | 365 Vues
Mechanisms of Transcriptional Repression.

E N D
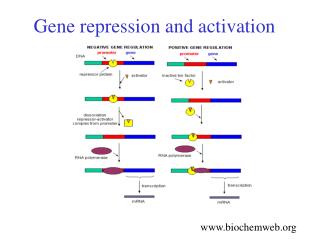
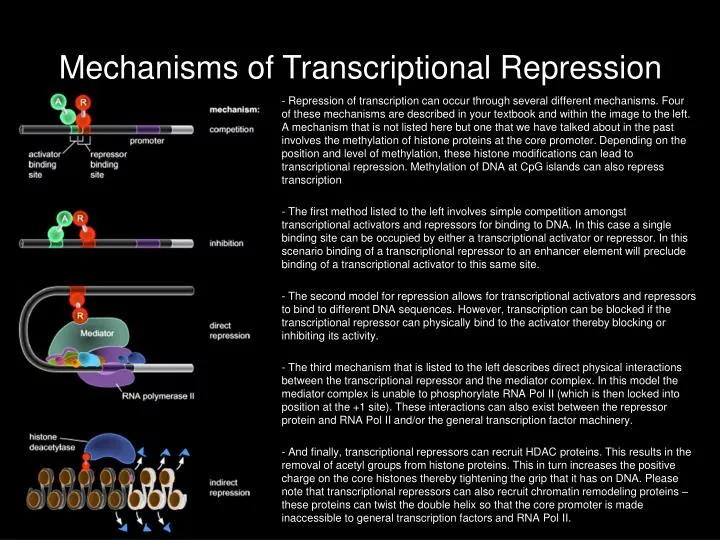
Mechanisms of Transcriptional Repression - Repression of transcription can occur through several different mechanisms. Four of these mechanisms are described in your textbook and within the image to the left. A mechanism that is not listed here but one that we have talked about in the past involves the methylation of histone proteins at the core promoter. Depending on the position and level of methylation, these histone modifications can lead to transcriptional repression. Methylation of DNA at CpG islands can also repress transcription - The first method listed to the left involves simple competition amongst transcriptional activators and repressors for binding to DNA. In this case a single binding site can be occupied by either a transcriptional activator or repressor. In this scenario binding of a transcriptional repressor to an enhancer element will preclude binding of a transcriptional activator to this same site. - The second model for repression allows for transcriptional activators and repressors to bind to different DNA sequences. However, transcription can be blocked if the transcriptional repressor can physically bind to the activator thereby blocking or inhibiting its activity. - The third mechanism that is listed to the left describes direct physical interactions between the transcriptional repressor and the mediator complex. In this model the mediator complex is unable to phosphorylate RNA Pol II (which is then locked into position at the +1 site). These interactions can also exist between the repressor protein and RNA Pol II and/or the general transcription factor machinery. - And finally, transcriptional repressors can recruit HDAC proteins. This results in the removal of acetyl groups from histone proteins. This in turn increases the positive charge on the core histones thereby tightening the grip that it has on DNA. Please note that transcriptional repressors can also recruit chromatin remodeling proteins – these proteins can twist the double helix so that the core promoter is made inaccessible to general transcription factors and RNA Pol II.
Rhodopsin: Capturing Photons of Light - Rhodopsin, a molecule comprised of the opsin protein and the retinal chromophore, is responsible for capturing photons of light and initiating the phototransduction cascade. The end result is a change in the voltage difference across the plasma membrane of the photoreceptor neuron. - The opsin protein is a member of a superfamily of G-protein coupled receptors. Each member of this family has seven transmembrane domains and is embedded within the plasma membrane. The genomes of seeing animals such as flies, mice and humans contain multiple opsin genes. The different opsin proteins have distinct spectral properties – in other words different rhodopsins capture light in different wavelengths. - Over the years it has been shown that a wide range of organisms including humans, elephants, mice, some Old and New World monkeys and even fruit flies can see in color. - In order to see in color photoreceptor neurons must not only express color sensitive opsins but individual photoreceptors must express one, and only one, opsin gene. The non-overlapping pattern of opsin gene expression is then used by the brain to compare inputs from the eye and see the world in color. In this lecture we will examine one mechanism for ensuring the correct opsin expression pattern.
The Drosophila Compound EyeSurface Topology and Structure of the Inner Photoreceptor Layer - As we have discussed before, the Drosophila compound eye is comprised of approximately 800 unit eyes that are also called ommatidia. Each ommatidium receives light from about 2 degrees of space. However, the fly does not see 800 different images. The images from each ommatidium are integrated within the brain and the fly actually sees a single image. - Each ommatidium contains eight photoreceptors numbered R1 thru R8. The photoreceptor neurons are homologous to the mammalian rod and cone photoreceptors. Each photoreceptor contains an organelle called the rhabdomere – all of the phototransduction machinery including the rhodopsin proteins are localized here . The rhabdomere is the equivalent to the rod and cone outer segments in the mammalian photoreceptor. - Six different rhodopsin proteins are encoded by genes within the fly genome (Rh1-Rh6). The Rh1 gene is expressed in photoreceptors R1 thru R6. The Rh2 gene is expressed in photoreceptors of the ocelli (which are not part of the compound eye). The Rh3 and Rh4 genes are expressed in subsets of R7 cells. And finally, the Rh5 and Rh6 genes are expressed in subsets of R8 cells.
Rhodopsin Genes in Drosophila Charles Zuker Steve Britt Karl Fryxell - The insect retina is a simple nervous system with a stereotyped developmental pattern and a limited number of cell types. In particular the fruit fly, Drosophila melanogaster, has been studied for over 100 years and this has allowed for the accumulation of numerous resources including a fully sequenced genome and large collections of cDNAs, cell lines genomic clones and antibodies. These features has made the fruit fly an ideal system for studying vision. - Charles Zuker and Joseph O’Tousa cloned the first invertebrate rhodopsin (Rh1) – this rhodopsin absorbs light in the orange spectrum and essentially used as a motion detector. - Charles Zuker, Karl Fryxell and Craig Montell went on to clone the Rh2, Rh3 and Rh4 rhodopsin genes – these absorb light in the ultraviolet spectrum. - Claude Desplan and Steve Britt cloned the Rh5 and Rh6 rhodopsins– these absorb blue and green light. They have shown that flies can see in color, a feature that was once thought to be restricted to a limited number of primates. - Flies have a color rhodopsin mosaic just like the human retina. Joseph O’Tousa Craig Montell Claude Desplan
Pattern of Rhodopsin Expression - The Rh1 protein is expressed in the six outer photoreceptors (R1-R6). It absorbs light in the orange spectrum and is required for motion detection. - The Rh2 protein is not expressed in the photoreceptors of the compound eye. Instead it is found within photoreceptors of three simple eyes (called ocelli) that sit at the vertex of the fly head - The Rh3 and Rh4 genes are expressed in non-overlapping sets of R7 cells. About 70% of ommatidia will have an R7 cell that expresses Rh3 while the remaining 30% of ommatidia will have an R7 cell that contains the Rh4 protein. These rhodopsins capture light in the ultraviolet spectrum. - The Rh5 and Rh6 genes are expressed in non-overlapping sets of R8 cells. About 70% of ommatidia will have an R8 cell that expresses Rh5 while the remaining 30% of ommatidia will have an R8 cell that contains the Rh6 protein. - If an ommatidium contains an R7 cell that expresses Rh3 then the R8 cell will express Rh5. On the other hand if an ommatidium contains an R7 cell that expresses Rh4 then the R8 cell will express Rh6. Rh1 Rh3 Rh4 4 3 4 3 4 3 5 2 5 2 5 2 8 8 8 1 1 6 1 6 6 7 7 7 4 3 4 3 5 2 5 2 8 8 1 1 6 6 7 7 Rh5 Rh6
Coordination of Rhodopsin Expression Steve Britt Claude Desplan - Steve Britt and Claude Desplan cloned the Rh5 and Rh6 rhodopsin gene, demonstrated that they are color sensitive and showed that they are only expressed in subsets of R8 cells. They also demonstrated that rhodopsin expression within an ommatidium is coordinated. For example, a single ommatidium can contain one of two pairs of cells: (1) an Rh3 expressing R7 cell and an Rh5 expressing R8 cell or (2) an Rh4 expressing R7 cell and an Rh6 expressing R8 cell. - The color sensitive rhodopsin genes (Rh5 and Rh6) are restricted to the R8 cells. Of particular interest to the Britt and Desplan labs has been the mechanisms that restrict rhodopsin expression. In this lecture we will describe a mechanism by which the Rh5 and Rh6 genes are restricted to the R8 photoreceptor neuron. 4 3 4 3 5 2 5 2 8 8 1 6 1 6 7 7 Rh4 Rh6 Rh3 Rh5
Properties of the Rh5 and Rh6 Enhancer - In the images above you are looking at tangential sections of the retina. In this view you can see the R7 photoreceptors which lie within the distal half of the retina and the R8 photoreceptors which lie within the proximal half of the retina (panel c). As we have discussed before, expression of the Rh5 and Rh6 genes are restricted to the R8 photoreceptor neurons. This can be seen using antibodies against Rh5 (panels C and E) and Rh6 (panel D). Note that the Rh5 and Rh6 are not expressed in the R7 cell layer (panels C and D). - A 246bp fragment (-1 to -246) upstream of the Rh6 +1 site is sufficient to drive expression of the lacZ reporter in a pattern of R8 cells that is identical to that of the wild type Rh6 gene. This suggests that all of the information required for normal Rh6 expression is contained within this 246bp element (panel E). The authors then determined that a slightly smaller fragment (-1 to -202) not only drives expression in the R8 cells but there is some expression in the R7 layer. This ectopic expression is the result of deleting the bases -203 to -246 and suggests that a transcriptional repressor binds to this 42bp sub-fragment.
Analysis of the seq56 Subfragment - The upstream genomic sequence of the Rh5 and Rh6 genes from three different Drosophila species were analyzed for similarities. In particular the 42 bases that contain the putative site for the transcriptional repressor were analyzed. An analysis of the six enhancer elements indicated that there is in fact a common sequence (white box). Since it is a common sequence found in the Rh5 and Rh6 enhancers the subfragment is called “seq56”. The sequence appears to be an imperfect palindrome with two halves that are called “A” and “B”. - The wild type 246bp enhancer can drive expression of lacZ in just the R8 layer (panel C). Mutating the B site results in ectopic expression in the R7 layer (panel D). Similarly, mutating the A site also leads to ectopic expression in the R7 layer (panel E). This result suggests that both halves of seq56 (A and B) are both necessary to prevent expression of Rh5 and Rh6 in the R7 cells.
Seq56 is Sufficient to Inhibit Transcription in the R7 Cell Layer - In the previous slide we determined that seq56 is necessary for the repression of Rh5 and Rh6 in the R7 cells. Here we will see if seq56 is sufficient on its own to repress transcription. The Rh3 gene is expressed in a subset of R7 cells and with the exception of a small subset of ommatidia at the dorsal rim Rh3 is only in the R7 layer. Studies from Gerald Rubin’s lab had identified an enhancer element that can drive expression of lacZ in the normal Rh3 expression pattern (panel F). If you replace the normal control elements with seq56 from the Rh6 enhancer you will abolish expression of lacZ in the the R7 layer (panel G). If you now take this construct and mutate the B half site you will restore expression of the reporter to the R7 cell layer (panel H). A similar result is seen if you mutate the A half site (not shown). These data suggest that seq56 is sufficient to inhibit transcription. The mechanism likely involves the binding of a transcriptional repressor to seq56. This repressor is likely to be expressed exclusively within the R7 photoreceptor neurons.
The Transcriptional Repressor Prospero Binds to seq56 within the R7 cell - Since Rh5 and Rh6 are repressed in the R7 layer the candidate transcriptional repressor that binds to seq56 most likely will be expressed exclusively in the R7 cell. A search for such a transcription factor identified the Prospero protein. It is only expressed in the R7 photoreceptor (panel A). In contrast, the Spalt transcription factor (Sal) is expressed in both the R7 and R8 layer (panel B). - Using an EMSA assay Prospero was shown to bind to seq56 (panel G blue box). Mutating the B half site abrogates binding of Prospero to seq56 (panel G red box) suggesting that Prospero binds to this site. Mutating the A half site has no effect on Prospero binding (panel G green box) indicating that Prospero does not bind to the A half site – it only binds to the B half site. - Note that in the reporter assays both the A and B half sites were both necessary for repression of Rh5 and Rh6. However, Prospero only binds to the B half site. This implies that a second transcriptional repressor binds to the A half site.
Molecular Biology Study Questions - How did the evolutionary analysis help in locating seq56? - What do the terms necessary and sufficient mean? - How do we know that a second transcriptional repressor is bound to seq56 in addition to Prospero? - How do we know the location of Prospero binding within seq56? - What would happen if seq56 was inserted into the Rh1 enhancer? - What would happen if Prospero was ectopically expressed in the R8 photoreceptor neurons? - What would happen if Prospero was ectopically expressed in the R1-R6 photoreceptors?
Preview of Upcoming Lecture Topics to be Covered Next Time Expression Profiling Using DNA Microarrays Dorsal-Ventral Patterning and Gene Expression Analysis • Textbook Chapter • Chapter 7pg168-171 Weekly Article(s) “Death Breath “Getting Close and Personal” “Regulating Evolution”