Download

1 / 57
590 likes | 783 Vues
Genomic Conflict and DNA Sequence Variation. Marcy K. Uyenoyama Department of Biology Duke University. Population genetics Historically model-rich Present need: model-based interpretation of observed patterns of genomic variation What are hallmarks of each model?

E N D
Genomic Conflict and DNA Sequence Variation Marcy K. Uyenoyama Department of Biology Duke University
Population genetics Historically model-rich Present need: model-based interpretation of observed patterns of genomic variation What are hallmarks of each model? Self-incompatibility systems in plants Recognizing genomic conflict due to sexual antagonism Overview
Neutral evolution Pure neutrality: distribution of offspring number is independent of any trait in parent Demographic history: deme founding, gene flow Purifying selection: maintain functioning state against random deleterious mutations Selection Balancing selection: maintenance of different forms Selective sweeps: substitution of most fit for less fit Canonical models
How do we know it when we see it? Patterns evident in genome variation Model selection Choosing among a small number of canonical models for any particular system Hallmarks of evolution
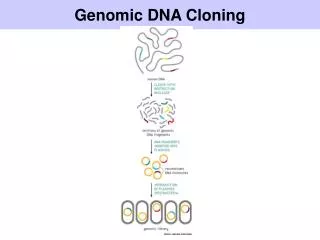
A random sample of genes Observed Sample Ancestral sequence
Allele and mutation spectra Site frequency spectrum Number of mutations Multiplicity a = {a1 = 6, a3= 1, a5= 1, a6= 1}, for aithe number of alleles with multiplicity i
The neutral coalescent Sample root from stationary distribution of P,mutation transition matrix and bifurcate • After an interval choose a lineage at random • Replace it by two identical copies with probability • Mutate it according to P with probability
Evolutionary rates • Events on level k • Bifurcation at rate Mutation at rate • Population parameters: ratios of rates • Next event is a bifurcation/coalescence with probability
Allele and mutation spectra Site frequency spectrum Number of mutations Multiplicity a = {a1 = 6, a3= 1, a5= 1, a6= 1}, for aithe number of alleles with multiplicity i
Mutation Novel allelic types formed at rate u per gene per generation Reproduction Frequency of allele i in the parental population: pi Multinomial sampling of N genes to form the offspring To find: probability of the sample of n genes (n1, n2, …, nk) or (a1, a2, …, an) for k the number of distinct haplotypes (alleles)ni the number of replicates of allele i ai the number of alleles with i replicates Infinite-alleles model
Ewens sampling formula a = (a1, a2, …, an), for ai the number of alleles represented by i replicates in a sample of size n θ = 2Nu, for N the effective number of genes and u the per-locus, per-generation rate of mutation Ewens (1972, Theoretical Population Biology)
Allele and mutation spectra Site frequency spectrum Number of mutations Multiplicity a = {a1 = 6, a3= 1, a5= 1, a6= 1}, for aithe number of alleles with multiplicity i
Population genomics About 750 accessions isolated from natural populations worldwide Summary statistics for sample of 19 entire genomes http://www.arabidopsis.org
Arabidopsis SNP spectra Minor allele counts 2 3 4 5 6 7 8 Site frequency spectra differ among functional classes Kim et al. (2008 Nature Genetics. 39: 1151)
Biallelic sample of size m Multiplicities i and (m – i ) ESF conditioned on two alleles independent of θ!
Ewens sampling formula a = (a1, a2, …, an), for ai the number of alleles represented by i replicates in a sample of size n θ = 2Nu, for N the effective number of genes and u the per-locus, per-generation rate of mutation Ewens (1972, Theoretical Population Biology)
Actual site frequency spectra • Excess of rare and common types, deficiency of intermediate types • Data from NIEHS Environmental Genome Project • Direct resequencing of loci considered environmentally-sensitive • Global representation of ethnicities Hernandez, Williamson, and Bustamante (2007)
Braverman et al. (1995) Spectrum shape • Signature of expansion? • Expansions maintain more rare mutations • Signature of selective sweep? • Neutral variants experience selection asa population bottleneck Black: constant population size Grey: recent expansion from small population size
Arabidopsis SNP spectra Minor allele counts 2 3 4 5 6 7 8 Site frequency spectra differ among functional classes Kim et al. (2008 Nature Genetics. 39: 1151)
Modelling a SNP data set Nordborg (2001 Handbook of Statistical Genetics) • Single segregating mutation in the sample genealogy • Conditional on exactly one segregating site, determine the distribution of the size (number of descendants) of the branch on which the mutation occurs • Exactly two alleles in the sample • Conditional on two haplotypes, bearing any number of segregating sites, determine the distribution of numbers of the two alleles
Two alleles One segregating site Conditioning
Single segregating site in a sample of size m Multiplicity i Multiplicity conditioned on a SNP dependent on θ! Ganapathy and Uyenoyama (2009 Theoretical Population Biology)
Arabidopsis SNP spectra Minor allele counts 2 3 4 5 6 7 8 Site frequency spectra differ among functional classes Kim et al. (2008 Nature Genetics. 39: 1151)
Population genetics Historically model-rich Present need: model-based interpretation of observed patterns of genomic variation What are hallmarks of each model? Self-incompatibility systems in plants Recognizing genomic conflict due to sexual antagonism Overview
Phenotypes Multiple genes generally influence a given phenotype Conflict Target trait value differs among genes that control phenotype Sexual antagonism Male and female function collaborate in reproduction Genes influencing each function may come into conflict Genomic conflict
Mating type regions as a battleground S-locus controls self-incompatibility in flowering plants How does sexual antagonism affect the pattern of molecular-level variation within the S-locus? What are hallmarks of conflict? Develop a basis for inference Model-based approach to the analysis of genetic variation Conflict and genomic variation
Flower development • Basic perfect flower includes both male and female components • Fertilization • Pollen grains deposited on stigma germinate and pollen tubes grow down style to the ovary Mariana Ruiz http://commons.wikimedia.org/wiki/File:Mature_flower_diagram.svg
Mariana Ruiz http://commons.wikimedia.org/wiki/File:Mature_flower_diagram.svg • Gametophytic SI (GSI) • Specificity expressed by individual pollen grain or tube determined by own S-allele • Pollen rejection • Growth of pollen tube arrested in style Norbert Holstein http://commons.wikimedia.org/wiki/File:Gametophytic_self-incompatibility.png
Mariana Ruiz http://commons.wikimedia.org/wiki/File:Mature_flower_diagram.svg • Sporophytic SI (SSI) • Specificity expressed by individual pollen grain or tube determined by the S-locus genotype of its parent • Pollen rejection • Germination of pollen grain may be arrested at stigma surface Norbert Holstein http://commons.wikimedia.org/wiki/File:Sporophytic_self-incompatibility.png
Mariana Ruiz http://commons.wikimedia.org/wiki/File:Mature_flower_diagram.svg Norbert Holstein http://commons.wikimedia.org/wiki/File:Gametophytic_self-incompatibility.png Pistil (A) component: rejection ofrecognized specificities Pollen (B) component: declaration ofspecificity An Bn Sn Norbert Holstein http://commons.wikimedia.org/wiki/File:Sporophytic_self-incompatibility.png
Mating type regions Uyenoyama (2005)
Human Y chromosome Skaletsky et al. (2003 Nature 423: 825) • Non-recombining male-specific Y (MSY) • Euchromatic region ~ 23 MB • Differences between two random Ys every 3 – 4 KB • Mammalian sex determinant SRY • Y-linked regulator of transcription of many male-specific Y-linked genes
Mating type regions • Linkage between pistil (A) and pollen (B)components is essential to SI function • Pollen: declaration of specificity • Pistil: rejection of recognized specificities Uyenoyama (2005)
Pollen component Pistil component Brassica S-locus Natural populations often contain 30 – 50 S-alleles Nasrallah (2000 Curr. Opin. Plant Biol.)
Ubiquitin tags proteins for degradation • Style: S-RNase disrupts pollen tube growth • Upon entering a pollen tube, S-RNases initially sequestered in a vacuole • In incompatible crosses, vacuole breaks down, releasing S-RNases into cytoplasm of pollen tube • Pollen: SLF (S-locus F-box) • Mediator of ubiquitinylation (attachment of ubiquitin) • Disables all S-RNases except those of the same specificity Vierstra (2009, Nature Reviews Molecular Cell Biology)
Pistil: why reject fertilization? Screening of potential mates may improve offspring quality Cost under incomplete reproductive compensation: ovules may go unfertilized Pollen: why provoke rejection? Self-rejection may improve quality of own ovules Rejection by other plants reduces siring success Hide behind another S-specificity in sporophytic SI? Decline to declare S-specificity altogether? Sexual antagonism
GSI model • Basic discrete time recursion • Symmetries in genotype and allele frequencies • Model change in frequency of focal allele i, assuming all other alleles in equal frequency Wright (1937, Genetics)
Diffusion approximation • Change in allele frequency • Diffusion equation coefficients • holds for large population size (N) and u (rate of mutation to new S-alleles) of order 1/N Wright (1937, Genetics)
Number of S-alleles Frequency in population Wright’s diffusion model • Diffusion with jumps • Turnover rate
Expansion of time scale under balancing selection • High rate of invasion of rare alleles • Promotes invasion of new and retention of rare types • Maintains high numbers of alleles • Genealogical relationships • Tree shape similar under symmetric balancing selection and neutrality • Greatly expanded time scale Takahata (1993, Mechanisms of Molecular Evolution)
S-allele turnover • Quasi-equilibrium of S-alleles Invasion of new, rare S-alleles balanced by extinction of common S-alleles • Expansion of time scale Rate of divergence among S-allele classes similar to rate among neutral lineages, but in a population of size fN:
Gametophytic SI models • Basic discrete time recursion • Diffusion approximation • Parameters: • Effective population size (N) • Rate of mutation to new S-specificities (u)
Simulation results • Stationary distribution of allele frequency • Most time spent close to deterministic equilibrium (1/n) or in boundary layer close to extinction • Number of S-alleles • Analytical expectation for number of common S-alleles Vallejo-Marín and Uyenoyama (2008)
Mariana Ruiz http://commons.wikimedia.org/wiki/File:Mature_flower_diagram.svg Norbert Holstein http://commons.wikimedia.org/wiki/File:Gametophytic_self-incompatibility.png Pistil (A) component: rejection ofrecognized specificities Pollen (B) component: declaration ofspecificity An Bn Sn Norbert Holstein http://commons.wikimedia.org/wiki/File:Sporophytic_self-incompatibility.png
Pollen specificity in GSI • Each pollen expresses its own specificity • Rarer specificities are incompatible with fewer plants • Incompatible matings • For n S-alleles in equal frequencies, a pollen type is incompatible with a proportion 2/n of all plants Norbert Holstein http://commons.wikimedia.org/wiki/File:Gametophytic_self-incompatibility.png
Pistil: why reject fertilization? Screening of potential mates may improve offspring quality Cost under incomplete reproductive compensation: ovules may go unfertilized Pollen: why provoke rejection? Self-rejection may improve quality of own ovules Rejection by other plants reduces siring success Hide behind another S-specificity in sporophytic SI? Decline to declare S-specificity altogether? Sexual antagonism
Relative viability of inbred offspring (s ) An+1 Bn Self-pollen fraction (s) Fate of style-part mutant Full SC Polymorphism Sa Full SI
An Bn+1 Fate of pollen-part mutant Full SC Relative viability of inbred offspring (s ) Sb Disruption Polymorphism Full SI Self-pollen fraction (s) Uyenoyama, Zhang, and Newbigin (2001)
An Bn+1 An+1 An+1 Bn Bn+1 Direction of pollen flow An Bn Sn Sb Sa Sn+1 Uyenoyama, Zhang, and Newbigin (2001)
An Bn+1 An+1 An+1 Bn Bn+1 An Bn TURN OFF Partial breakdown of SIby pollen disablement Sn Evolutionarily unlikely Sb Sa TURN ON Restoration of SIby stylar recognition Evolutionarily unlikely Sn+1 Uyenoyama, Zhang, and Newbigin (2001)