Download

1 / 37
370 likes | 499 Vues
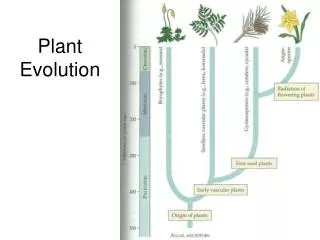
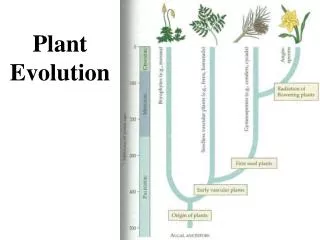
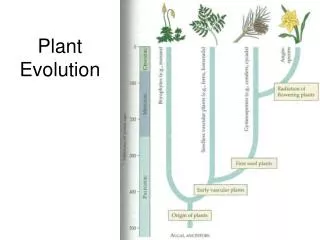
Evolution of plant-herbivore relationships. Photo G. Weiblen. Cospeciation resulting in congruent phylogenies. Host-shift speciation resulting in congruent phylogenies, but with shorter branches in the parasite lineages. de Vienne et al. New Phytologist (2013) doi: 10.1111/nph.12150.

E N D
Evolution of plant-herbivore relationships Photo G. Weiblen
Cospeciation resulting in congruent phylogenies. Host-shift speciation resulting in congruent phylogenies, but with shorter branches in the parasite lineages de Vienne et al. New Phytologist (2013) doi: 10.1111/nph.12150
Host-shift speciations, resulting in incongruent phylogenies. de Vienne et al. New Phytologist (2013) doi: 10.1111/nph.12150
Evolution of insect-plant associations: by descent or colonization plant insect Association by descent Association by colonisation Sequential evolution: the evolution of herbivorous insects follows the evolution of plants while the plant evolution is not affected by herbivores. Coevolution: the evolution of plant lineages influences the evolution of herbivore lineages and vice versa
Coevolution: reciprocal evolutionary change in interacting species Escape-and-radiate coevolution 1. Plants evolve by chance (via mutation, recombination) a new toxin/deterrent. 2. New chemical leads to protection from herbivores. 3. Protected plants enter a new adaptive zone, in which they are free to radiate. 4. Herbivores evolve (via mutation, recombination) ways to deal with new toxin. 5. Herbivores enter a new adaptive zone and are free to radiate. 6. The cycle is repeated.
Escape and radiation concept: plants develop new defence [yellow, orange], their speciation rate increases, herbivores develop counter-defence and colonize them Futuyma & Agrawal, 2009, PNAS 106: 18054–18061
Evolution of plant defense traits and herbivore specificity If most herbivores are generalists, and only a subset of the plant species pool can defend or tolerate the dominant enemies, then plant species composition will shift to become dominated by those species that share these defence and tolerance traits. In this figure, green squares, red stars and orange circles represent different defence traits that confer tolerance of herbivory in plants within a community, and thin lines indicate a species has been eliminated from a community by the herbivore. If defence traits are conserved (a), heavy herbivore pressure will drive phylogenetic clustering within the community. For example, large mammalian herbivores consume a wide variety of plants, yet grasses are able to tolerate high herbivory pressure and in the presence of these large herbivores, quickly dominate communities. If herbivores are excluded, plant composition changes, and trees or forbs can take over (McNaughton 1985, Pringle et al. 2007). However, if such traits that confer tolerance or defence are convergent, generalist enemies will drive the phylogenetic community structure towards overdispersion (b). If specialists exert a large proportion effect on plant fitness within a community, this will result in strong patterns of density dependence (Janzen 1970, Connell 1971). This should increase local diversity by favouring rare species which can escape their natural enemies more often than more abundant species. Furthermore, if related plants have qualitatively similar defence strategies (trait conservatism) (c), strong Janzen–Connell regulation in a community could limit the co-occurrence of closely related species and promote the co-occurrence of distantly related species at neighbourhood scales, causing community phylogenetic overdispersion (Webb et al. 2006). In this figure, specialist enemies can eat only plants from the pool that have similar defence traits, similar to Becerra (1997). (d) If plants defence traits are convergent, however, Janzen–Connell regulation by specialist enemies will promote random patterns in plant communit phhylogenetic structure. Cavender-Bares et al. Ecology Letters, (2009) 12: 693–715
Gall wasps (Cynipidae): second largest radiation of gallers, 1300 spp. Most of species on woody plants, particularly oaks (a), species on herbaceous plants mostly on Asteraceae (b) Phylogenetic conservatism in the position of galls on host plants
Cynipid gallers Andricus on oaks in Europe: evolutionary conservatism in gall type
inquilines do not induce their own galls, feed inside galls of other spp. Host plants of gallers
Phyllobrotica beetles and Scutellaria hosts: parallel (or sequential?) evolution S. galericulata P. quadrimaculata Farrell & Mittter 1990
Tetraopes beetles and Asclepias hosts: an example of escape-and-radiate coevolution? Farrell & Mitter 1998
Blepharida beetles on Bursera plants: secondary chemistry explains multiple host colonizations different secondary chemistry marked by different colour Host selection by beetles can be better explained by plant similarity in secondary metabolites than by plant phylogeny Bursera phylogeny Bursera phylogeny does not correspond with phylogeny of its beetles Becerra 1997
Asclepias defense strategies: life history traits and phylogeny A. californica A. exaltata A. asperula Phenogram recognizing three defense strategies based on 7 traits: Distribution of defense strategies on Asclepias phylogeny A: soft leaves, many trichomes, high latex B: tough leaves, low water content, medium latex C: soft leaves, low latex, high cardenolides Agrawal & Fishbein, Ecology, 87 Suppl., 2006, S132–S149
Asclepias defense strategies: phylogenetically unstable Agrawal & Fishbein, Ecology, 87 Suppl., 2006, S132–S149
Phylogenetic and physiological response may not be necessarily the same Asclepias syriaca • Latex and trichomes: • no correlation within species (Asclepias syriaca) • a positive correlation across 24 species of Asclepias Agrawal and Fishbein 2006
Herbivores feeding on latex-rich Asclepiadaceae - Apocynaceae colonize preferably other latex plant lineages Number of colonization of plants from various orders by herbivores feeding on latex-rich Asclepiadaceae - Apocynaceae plants: insects retain their taste for latex
Many herbivore lineages retain broad preferences for certain plant lineages
Many herbivores have polyphyletic diets generated by multiple host colonisations Herbivore species Plant species
Swiss Alps: 231 most abundant plant species vs. all butterfly species Pellissier et al. 2013, Ecology Letters
Where to find genuine plant - insect coevolution? • The plant - insect mutualisms where • the insect pollinates flowers • then oviposits to some of them so that the larval survival depends on successful pollination Ficus - Agaonidae wasps Yucca - Tegeticula moths (Yponomeutidae) Trollius - Chiastocheta (Anthomyiidae) flies Glochidium - Epicephala (Gracillariidae) moths
male female Fig inflorescence: flowers are hidden inside
Ceratosolen wasps: Oviposition to fig flowers through stylus
Epicephala sp. (Gracillariidae) and Glochidium (Euphorbiaceae) Kato et al. 2003. PNAS 100:5264
Is narrow host specialization an evolutionary dead-end? Specialized Chrysomela beetles dependent for anti-predator defence on metabolites from their hosts develop a new chemical defense butyric acids salicylaldehyde from salicin monoterpene iridoids Termonia et al. 2001
Mass extinction (Cretaceous-Tertiary boundary): specialists die first Labandeira et al. 2002 PNAS 99:2061
angiosperms conifers Cycadales 1 - 5: colonizations of angiosperms Beetles: high species diversity associated with feeding on angiosperms Farrell 1998
Host specificity: is narrow specialization determined by speciation dynamics? Transition from a generalist to a specialist is more likely than reverse transition Nosil 2002
No. of mouthpart classes No. of insect families While number of insect families is steadily increasing through evolution, there has not been any major inovation in mouthparts dor almost 100 million years: has everything been already invented?
Extinction of large mammalian herbivores in past 130,000 years: what are ecological consequences?
What were ecological roles or recently extinct megafauna? Janzen & Martin (1982) NEOTROPICAL ANACHRONISMS: The Fruits the Gomphoteres Ate Large recently extinct fauna, such as gomphoteres in S. America, could be important consumers and dispersal agents of large fruits Crescentia alata Enterologium cyclocarpum Similar role played by forest elephants in Africa
Leaf-cutting ants: an accident of evolution?
before Atta nest after