RNA Processing
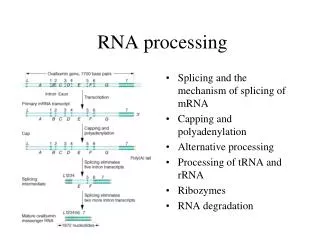
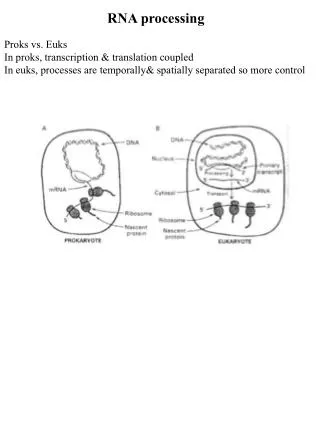
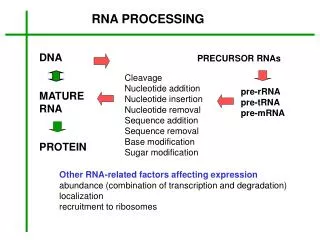
RNA Processing. Processing and Export of RNA. Overview. In eukaryotes, the growing functional complexity and cellular compartmentalization requires that RNA be processed for long term storage and transport through nuclear membrane. rRNA porcessing tRNA porcessing mRNA porcessing 5’m caps


RNA Processing
E N D
Presentation Transcript
Processing and Export of RNA Overview In eukaryotes, the growing functional complexity and cellular compartmentalization requires that RNA be processed for long term storage and transport through nuclear membrane. • rRNA porcessing • tRNA porcessing • mRNA porcessing • 5’m caps • 3’polyA tail • Splicing • Spliceosomes • Self splicing RNAs • Mutations in splicing • Alternative splicing • RNA editing • RNA export
Processing of rRNA Transcripts • rRNA constitutes 95% of the total RNA in the cell. • Multiple copies of rRNA genes are tendamly arranged, separated by spacer regions. • The pattern of alternating transcribed and non-transcribed region is readily obvious.
Ribosomal RNA Processing • The rRNAs are named according to their "S" values, • which refer to their rate of sedimentation in an • ultra-centrifuge. • The larger the S value, the larger the rRNA. • 45S precursor rRNA is chemically modified and • enzymatically cleaved in to smaller S fragments. • Two types of chemical modifications are added: • Psueodouridine • 2’O methyl ribose. • The spacer sequences between 18S, 5.8S and 28S • rRNA are discarded and degraded. • .
tRNA Processing • A tRNA molecule consists of 70-80 nucleotides. • Some nucleotides in tRNA are modified to: • dihydrouridine (D), • pseudouridine (Y), • inosine (I), • Inosine plays an important role in codon recognition. • In addition to these modifications, a few nucleosides are methylated. • The major role of tRNA is to translate mRNA sequence into amino acid sequence.
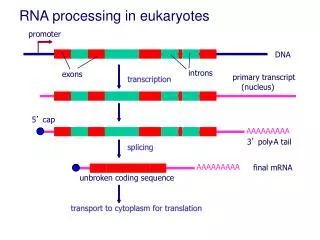
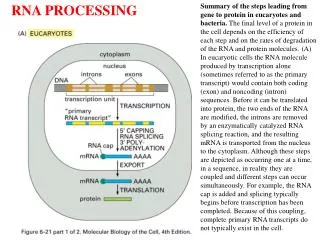
Structure of Mature Eukaryotic mRNA • All mRNAs have a 5’ cap • All mRNA (except histone) have a 3’ poly(A) tail. • the 5’ cap and 3’ poly(A) tail prevent mRNA degradation.
Messenger RNA Processing • Overview: • 5’-Capping • 3’-polyadenylation • Splicing • Splice site recognition • Spliceosomes • Self splicing RNAs • Mutations that disrupt splicing • Alternative splicing
5’ Capping • Capping occurs co-transcriptionally. • 7-methly guanosine (7mG) cap is added to the 5’ end. • Guanyltransferase forms a 5'-5' linkage between • 7-methyl guanosine and 5’-PO4 of the first base. • This methyl group is called Cap0. • O-methyl transferase then adds a methyl group to • the 2’-OH of the first two original bases in RNA. • These methyl groups are referred to as CAP1 and CAP2. • Methyl groups protect mRNA from degradation.
5’-Methylated Caps in mRNA • Methylated bases in caps 0 and 1. • m7G-5'-p-p-p-5'-Adenosine-3'-p-etc • O-Methylated sugars in caps 1 and 2.
3’-Polyadenylation of mRNA Pimary RNA transcript is cleaved and modified by RNA specific proteins. Cleavage and polyA signal is located ~ 30 residues upstream of a GU rich termination site.
3’-Poly-Adenylation of mRNA • An Endonuclease cleaves the primary transcript at CA, • ~10-30 nucleotides downstream of the AAUAAA. • Poly-A polymerase then adds ~ 200 adenosine residues. • Poly-A tail is associated with Poly-A binding proteins. • Poly-A tails stabilize mRNA.
RNA can fold into specific structures (A) Diagram of a folded RNA structure showing only conventional base-pair interactions; (B) structure with both conventional (red) and nonconventional (green) base-pair interactions; (C) structure of an actual RNA. Each conventional base-pair interaction is indicated by a "rung" in the double helix. Bases in other configurations are indicated by broken rungs.
Splicing of mRNAs • Coding sequences in eukaryotic RNA is frequently interrupted by non-coding regions. • Removal of intervening sequences or Introns (grey) from pre-mRNA transcripts. • Joining of the expressed sequences or Exons (color) to formmature mRNA transcripts. • Splicing occurs in the nucleus before transport to the cytoplasm. • cDNA contain only exons. • Genomic DNAs contain both exons and introns. • Incidence of introns increases with complexity in organisms.
Variation in intron and exon lengths in the human, worm, and fly genomes A) Size distribution of exons. (B) Size distribution of introns. Note that exon length is much more uniform than intron length. (Adapted from International Human Genome Sequencing Consortium, Nature 409:860 921, 2001.)
Heteronuclear Ribonucleoproteins (hnRNPs) Self Hybridization of RNA molecules in vitro is prevented by hnRNP proteins. The presence of complex secondary structures within RNA molecules inhibits hybridization between long complementary sequences in separate molecules. Association of hnRNP proteins with RNA is thought to prevent formation of RNA secondary structures, thereby facilitating base-pairing between different complementary molecules during splicing events. These proteins may have a similar function in vivo.
Splicing signals • Splice Junction Consensus Sequence : • In over 60% of cases, the upstream exon sequence is (A/C)AG at the donor site, • and G at the downstream splice acceptor exon site. • 5’-Donor site: AG | GUAAGU, where GU is always at 5' end of intron • 3’-Acceptor site: (Py...Py) 12 NCAG| N, where AG is always at 3' end of intron. • 2’- Branch site: CU(A/G)A(C/U) where A is conserved in all organisms.
The consensus Splice Signal Sequences in an RNA molecule Only the three blocks of nucleotide sequences shown are required to remove an intron sequence; the rest of the intron can be occupied by any nucleotide. Here A, G, U, and C are the standard RNA nucleotides; R stands for either A or G; Y stands for either C or U. The A highlighted in red forms the branch point of the lariat produced by splicing. Only the GU at the start of the intron and the AG at its end are invariant nucleotides in the splicing consensus sequences. The remaining positions (even the branch point A) can be occupied by a variety of nucleotides, although the indicated nucleotides are preferred. The distances along the RNA between the three splicing consensus sequences are highly variable; however, the distance between the branch point and 3’ splice junction is typically much shorter than that between the 5’ splice junction and the branch point.
Splicing of Eukaryotic pre-mRNA 2-Step Trans-esterification: • Cleavage at the 5'-end (G) of the intron by attack of a specific 2'OH group (A), the branch site. • This forms a 2’-5’phosphodiester bond with the 5'-phosphate of the intron, creating a lariat structure. • Ligation of the 3'-OH on Exon 1 with the 5‘- phosphate of Exon 2 then releases the lariat (RNA) intron.
Splicing Branch Point The structure of the branch point in the lariat intermediate in which the adenylate residue is joined to three nucleotides by phosphodiester bonds. The new 2’-to-5’ linkage is shown in red, and the usual 3’-to-5’ linkages are shown in blue.
donor (5’) splice site branch site acceptor (3’) splice site G/ GU AAGU..................…A.......… YYYYYNY AG / G U1 U2 Spliceosome mediated Splicing • In most eukaryotes, splicing is mediated by a large ribonucleoprotein complex, • the spliceosome. • The spliceosome contains a specific set of U-rich small nuclear RNAs associated with • ribonucleoproteins or snRNPs (snurps). • Splicing snRNPs: • U1: 5'- site recognition • U2: branch site recognition • U4: forms base paired complex and acts with U6 • U5: 3'- junction binding of U4-U6 complex • U6: complex with U4 makes up the spliceosome transesterase. • The common spliceosome recognizes introns starting with 5'-GU and ending in AG-3‘
Pre-splicing Complex Interactions between pre-mRNA, U1 snRNA, and U2 snRNA early in the pre-splicing process. The 5’region of U1 snRNA initially base-pairs with nucleotides at the 5’end of the intron (blue) and 3’end of the 5’exon (dark red) of the pre-mRNA; U2 snRNA base-pairs with a sequence that includes the branch-point A, although this residue is not base-paired. Secondary structures in the snRNAs that are not altered during splicing are shown in diagrammatic line form. The purple rectangles represent sequences that bind snRNP proteins recognized by anti-Sm antibodies. For unknown reasons, antisera from patients with the autoimmune disease systemic lupus erythematosus (SLE) contain these antibodies. Such antisera have been useful in characterizing components of the splicing reaction.
snRNP Spliceosome • Pre-splicing complex is formed by U1 and U2. • Tri snRNP U5, U4:U6 is then recruited. • U4 is base paired with U6. • U5 then displaces U1 from the upstream exon. • U6 then base-pairs to U2, resulting in displacemant of U4. • Finally, U5 base pairs to exon 2 near the 3'- splice site on • the same stem loop that already holds Exon1. • This brings the 3'-OH of Exon 1 into close proximity to • 5'-p of exon 2. • Nucleophilic attack of the phosphodiester bond • completes the splicing process, releasing the intron as • a lariat carrying the various splicing factors.
Self Splicing Catalytic RNAs Lower eukaryotes (ribozymes) and mitochondrial RNAs can undergo self splicing without the help of other splicing factors. Group I introns Group II Introns
Secondary structures of group II self-splicing introns (a) and U snRNAs present in the spliceosome The first transesterification reaction is indicated by black arrows; the second reaction, by blue arrows. The branch-point A is boldfaced. The similarity in these structures suggests that the spliceosomal snRNAs evolved from group II introns, with the trans-acting snRNAs being functionally analogous to the corresponding domains in group II introns