Gene Expression Patterns in Dimorphic Sperm Cells of Plumbago zeylanica
This study investigates the gene expression profiles in the dimorphic sperm cells of Plumbago zeylanica, which play crucial roles in fertilization. We utilized mRNA isolated from each sperm cell type to construct cDNA libraries and characterized patterns using microarrays and qRT-PCR. Results reveal distinct expression levels in the Sua and Svn sperm cells, suggesting their differing fates during fertilization—where Sua predominantly fertilizes the egg cell and Svn forms the endosperm. The findings enhance our understanding of male gamete development in angiosperms.


Gene Expression Patterns in Dimorphic Sperm Cells of Plumbago zeylanica
E N D
Presentation Transcript
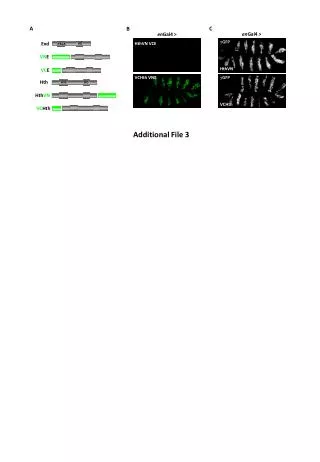
Male gamete development A B C A B C Meiosis Mitosis I Mitosis II A A Pollen D E F Micro- sporocyte D E F Generative cell Vegetative cell Sperm cells Tetrad Female gamete development Egg cell H G I G H I Germinated pollen Double fertilization Sperm Sua + Egg Zygote Embryo Sperm Svn + Central Cell Endosperm Central cell J L K K L J N O M M N O Sua Svn R Q P VN Q P R S U T S T X W V C In situ hybridization Whole mount In situ hybridization was undertaken to confirm patterns of transcription of selected clones in mature pollen of Plumbago zeylanica using digoxigenin RNA probes. Five transcripts shown below in Figure 7 illustrate three major patterns of gene expression in sperm cells: (1) nearly equal expression for clones SuaCon16 and SvnA3F16; (2) strongly upregulated in the Sua in ubiquitin E2 and SuaCon62; and (3) very strongly upregulated in the Svn in isopentenyl transferase. The latter two patterns were confirmed with RT-PCR (data not shown). Gene expression in the dimorphic sperm cells of the angiosperm, Plumbago zeylanicaXiaoping Gou, Tong Yuan, Xiaoping Wei and Scott D. Russell* Department of Botany and Microbiology, University of Oklahoma, Norman, OK 73019 USA ABSTRACT Mature pollen of the flowering plant Plumbago zeylanica contains dimorphic sperm cells that differ in organization and fate. Sperm cells are linked with the vegetative nucleus (VN) in pollen creating a polarized functional unit for co-transmission of DNA known as the “male germ unit” (MGU). In the MGU, one sperm cell (Svn) is physically associated with the VN, whereas the other linked sperm cell (Sua) is unassociated with the VN. Since the two sperm cells differ in their organelle complement, their fate can be traced and is known to be preferential, with the Sua fusing with the egg cell in >95% of cases (forming the zygote and embryo), whereas the Svn fuses with the central cell forming the nutritive endosperm during double fertilization. To determine whether gene expression differences relating to fertilization fate occur in the two sperm cells, we isolated mRNA from each sperm cell population, created PCR-cDNA libraries (both representative and subtracted) and characterized products using ESTs, microarrays, qRT-PCR and in situ hybridization. Large numbers of library members, yielded no hits (43%), or unclassified and hypothetical proteins (17%); this is similar to animal sperm cells in that a large percentage of mRNA products could not be functionally categorized (Dadoune et al. 2005). Expressional profiles based on products examined using qRT-PCR revealed that about half showed products that were uniquely or highly expressed only in sperm cells (screened against three pollen stages, stem, root, petal, sepal, and ovary. Sequences for nearly 2000 clones are posted at URL: http://www.genome.ou.edu/plumbago.html with 1522 in GenBank. To screen for differential expression in the two sperm cells, we made suppression subtraction hybridization (SSH) PCR-amplified libraries and spotted 2304 clones from each library onto glass slides for microarray screening, using hybridization with SSH and non-SSH cDNA targets. Many clones expressed differential expression patterns in the two sperm cell types and 24 were selected for qRT-PCR confirmation. From these candidates, we obtained several products expressed in only one sperm cell, corroborated by whole mount in situ hybridization. We also isolated sperm-specific promoters that display GFP and GUS activity in sperm cells of Arabidopsis. The most abundant product in the endosperm-targeted Svn sperm represents isopentenyl transferase homologs relating to synthesis of cytokinin—a plant hormone known to stimulate early endosperm activation. We suspect transmission of sperm mRNA during fertilization may be occurring in flowering plants, similar to that reported in animals (Ostermeier et al. 2004) and that this sperm mRNA may directly influence early post fusion activation events. RESULTS cDNA library construction and EST analysis The initial library was a PCR-based cDNA library constructed using the Smart cDNA library kit (Clontech) based on RNA isolated from 12,000 sperm cells of each morphotype to construct two size-selected (>400 bp) cDNA libraries. The titers of the unamplified libraries were 2.1 x 107 and 3.2 x 107 pfu/ml, for Sua and Svn, respectively. The libraries are of high quality: about 90% of the clones had inserts and over 95% of the inserts were at least 500 bp. About 2000 ESTs were sequenced in all. Initial sequencing of the libraries was conducted until products produced about 80% redundancy. The Sua collection included 583 unique EST sequences, forming 191 contigs and 392 singlets, of which 427 are distinct from Svn sequences. The Svn collection included 471 unique EST sequences, with 122 contigs and 349 singlets, of which 330 are different from Sua sequences. About 70 ESTs were found in both sperm cells and about 80 more had probable matches below E-60. Many products had no known homology (43.4% for Sua, 46.9% for Svn), another large group consisted of unclassified or hypothetical proteins (16.5% for Sua and 14.9% for Svn) and the remainder were categorized according to Gene Ontology functional assignments (http://www.geneontology.org/) Figure 3 provides an overview of ESTs for which functional assignments could be made and displays their similarity prior to assembly of overlapping ESTs into contigs. These results reflect that the two libraries are outwardly similar between the two cells. The number of Sua products appears to be somewhat more diverse, based on sequence numbers. Although this is not necessarily an accurate estimate of cDNA library members, there is an impression of greater diversity, which may correspond with its future fate as male partner in zygote activation and embryo form. Suppression subtraction hybridization (SSH) cDNA libraries to each sperm type resulted in markedly different profiles. SSH-cDNA results are categorized in Figure 4, reflecting markedly different profiles between the Sua and Svn, which appear to relate to their future female target. The Sua yielded more ESTs involved with transcription, translation, and post-translation modification. In contrast, the Svn (which forms endosperm) yielded more ESTs in biosynthesis and energy-related metabolic pathways. Figure 7. Whole mount In situ hybridizations of mature pollen labeled with digoxigenin antisense RNA probe in A-O and control sense probe in P-R. Whole mounts are shown in brightfield microscopy (columns 1 & 4), mixed brightfield/DAPI epifluorescence (cols. 2 & 5), and DAPI epifluorescence (cols. 3 & 6). A-F: Nearly equal expression between two sperm cells in clones SuaCon16 and SvnA3F16; G-L: Strong upregulation in Sua in clones of ubiquitin E2 and SuaCon62; M-O: Strong upregulation of isopentenyl transferase in Svn. P-R: Same as M-O with sense probe. Sua Sua Plumbago promoter isolation and expression Genes showing Sua- or Svn-based specificity were selected for the isolation of Plumbago promoters. Using sequences of identified full length cDNA clones, a set of nested primers of each gene were designed to isolate upstream genomic DNA sequences from a genomic DNA library. Promoter fragments were then fused with GFP or GUS and cloned into plant transformation binary vectors and transformed into Arabidopsis and tobacco using standard techniques. Promoters retained expression in MGU cells, including the generative cell of tobacco (Figure 8A-B) and the sperm cells of Arabidopsis (Figure 8C), displaying conserved activity despite a significant phylogenetic distance. Conservation of activity can also be retained between monocots and dicots, as shown by expression of the LGC1 promoter of lily (Xu et al. 1999), which has been successfully expressed in tobacco (Singh et al. 2003) and Arabidopsis (Gou and Russell, not shown). INTRODUCTION Flowering plants require two fertilization events to effect successful formation of fertile seed: (1) a fusion of one sperm cell with the egg cell, forming the zygote and subsequent embryo and (2) fusion of a second sperm cell with the central cell with its multiple polar nuclei resulting in formation of endosperm—a typically polyploid tissue unique to angiosperms that provides nutrition for the embryo. Flowering plants thus use both of the sperm cells delivered by the pollen tube to achieve seed formation in an event known as double fertilization. The different developmental fates of embryo and endosperm are believed to arise from the extreme dimorphism and presumed gene expressional differences between the egg and central cells. Potential expressional differences in the sperm cells, and their potential role in determining the fate of the sperm cells and their post-fertilization products, however, have not been addressed before. The flowering plant, Plumbago zeylanica, is a model species for the examination of sperm cell differences, as it has been the subject of continuing study and is known to display highly distinctive sperm cell types in which the fate of each sperm cell during fertilization has been described. Examining gene expressional differences between the two sperm cells may therefore reveal how sperm cell dimorphism arises in the parent generative cell, and how the two sperm cells may be targeted during double fertilization and influence early post-fertilization development. The relationship of the sperm cells is displayed below (Figure 1), reflecting the unique cell-within-a-cell relationship of sperm within pollen grains in tricellular plants. Sperm cells are non-motile, nutritionally and physically dependent on the pollen tube to deliver them to the female target cells during double fertilization. Svn Svn Figure 3. Functional categorization of assignable EST sequences of the Sua and Svn from putatively representative cDNA libraries. Although ESTs differed between the two cells, the distribution of functional categories was largely similar over most categories. The most conspicuous exceptions included upregulation of posttranslational modification in the Svn and transcription in the Sua. Figure 4. Functional categorization of assignable EST sequences of the Sua and Svn from SSH-cDNA libraries. The Sua showed upregulation of products involved with transcription, translation, post-translation modification and DNA replication, with downregulation of most energy-related metabolic pathways. In contrast, the Svn showed strong upregulation of biosynthesis and energy-related metabolic pathways. Figure 8. Activity of Plumbago promoter was documented by GUS or GFP labeling. A,B: Tobacco pollen showing activity of pA7::GUS in generative cell, viewed using DAPI fluorescence and brightfield microscopy. C: Arabidopsis pollen showing activity of pIPT::GFP in sperm cells, viewed using confocal microscopy. qRT-PCR assays of expression patterns in different tissues qRT-PCR was used to understand the extent to which sperm cell genes were expressed in other tissues, how many were essentially expressed exclusively in sperm cells and whether gene products were differentially expressed in the two sperm cells. To examine these patterns, sperm were compared with the following developmental stages of pollen: microspore, bicellular and mature pollen; the following sporophytic vegetative tissues: root, stem and leaf; and the following sporophytic reproductive organs: sepals, petals, ovary before and after pollination. Equal quantities of template were used for each experiment and the expression level of the Sua was set as 1. Clones were essentially randomly selected relative to expressional pattern. Of analyzed samples, about half show no appreciable expression outside of sperm cells. Figure 5 shows a gallery of products from the representative cDNA libraries. Figure 6 shows a gallery of products from the SSH-cDNA libraries. CONCLUSIONS Sperm cells in flowering plants display a rich and complex assemblage of mRNAs (Engel et al. 2003). In our study, we found about half are expressed differently in a variety of tissue types and another half are significantly upregulated or expressed only in cells of the male germ lineage—the generative and sperm cells—not even overlapping with pollen cytoplasm. The dimorphic sperm cell types produced complementary cDNA libraries that displayed reasonably similar frequencies of ESTs within assignable Gene Ontology functional categories. However, use of SSH-cDNA libraries for screening, combined with microarray and qRT-PCR, reveals numerous expressional differences. Postulating from Plumbago to Arabidopsis, it seems possible that numerous genes described through genomics that have been unsupported by ESTs to date may be expressed in sperm cells. In Plumbago zeylanica, the Sua sperm cell typically fertilizes the egg forming the zygote and embryo and the Svn typically fertilizes the central cell, forming the endosperm. Among the most highly expressed of Svn genes are those involved in cytokinin synthesis and various forms of cellular metabolism, which are seemingly inconsistent with sperm and pollen function, but strikingly consistent with the activity of endosperm; endosperm is largely controlled by cytokinin activity and is a nutritive tissue for the embryo. Similarly, Sua genes displayed enrichment of transcription-, translation- and post-translation-related transcripts that seemed more consistent with zygote activation than with sperm function. If plants transmit and express sperm mRNA in a manner consistent with that in humans, the influx of specific sperm transcripts may contribute to the divergence of developmental programs and thus sperm dimorphism could contribute to accelerated development of seed. The sperm cells have promoters corresponding to this behavior. Sperm mRNA of plants displays high frequency of ESTs with no known homology, putative and unknown proteins—more than many other known tissues. of which as many as 50% of the most commonly collected ESTs may correspond to sperm specific transcripts. Thus, sperm cells may be one of the least well described for gene transcription and expression. Figure 1. Origin of non-motile sperm cells within the pollen grain and tube, and fate of sperm cells during double fertilization in Plumbago. Sua • REFERENCES • Dadoune JP, Pawlak A, Alfonsi MF, Siffroi JP (2005) Identification of transcripts by macroarrays, RT–PCR and in situ hybridization in human ejaculate spermatozoa Mol Hum Reprod 11: 133-140 • Engel ML, Chaboud A, Dumas C and McCormick S. 2003. Sperm cells of Zea mays have a complex complement of mRNAs. Plant J, 34:697-707 • Ostermeier GC, Miller D, Huntriss JD, Diamond MP, Krawetz SA (2004) Reproductive biology: Delivering spermatozoan RNA to the oocyte. Nature 429: 154 • Russell, SD. 1984. Ultrastructure of the sperms of Plumbago zeylanica. 2. Quantitative cytology and three-dimensional organization. Planta 162:385-391 • Russell, SD. 1985. Preferential fertilization in Plumbago: ultrastructural evidence for gamete-level recognition in an angiosperm. Proc Natl Acad Sci USA, 82:6129-6132 • Singh M, Bhalla PL, Xu H, Singh MB. 2003. Isolation and characterization of a flowering plant male gametic cell-specific promoter. FEBS, 542:47-52 • Xu H, Swoboda I, Bhalla P L, and Singh M B. 1999. Male gametic cell-specific gene expression in flowering plants. Proc Natl Acad Sci USA, 96: 2554-2558 • Zhang Z, Xu H, Singh MB, Russell SD. 1998. Isolation and collection of two populations of viable sperm cells from the pollen of P. zeylanica. Zygote, 6: 295-298 B Svn Figure 5. Gallery of qRT-PCR results of selected cDNA library members prepared using ABI Prism 7000 SDS with expression level of Sua set to 1. Among sperm cell expressed products about half showed nearly equal expression between the two sperm cell types. The remainder had more products In which the Sua (which usually fuses with the egg cell) was more highly upregulated. Clones were selected essentially randomly relative to overall expressional pattern in tissues other than sperm cells. Of analyzed samples, about half show no or little appreciable expression except in sperm cells. Samples for each chart are (left to right): 1, Microspore; 2, Bicellular pollen; 3, Root; 4, Stem; 5, Leaf; 6, Mature pollen; 7, Sepal; 8, Petal; 9, Ovary before pollination; 10, Ovary after pollination; 11, Sua; 12, Svn. Figure 2. (A) Male germ unit in the pollen tube (PT) showing vegetative nucleus (VN) and its associated leading sperm (Svn) and the trailing sperm cell (Sua). (B) Organellar dimorphism in Plumbago zeylanica (n=11) is based on serial ultrastructural reconstruction of paired sperm cells in pollen. (C) Isolation of sperm cells used for obtaining mRNA from pure populations, showing freshly isolated sperm cells (upper left), sperm after 2 min (lower left), collection of only Sua sperm cells (center) and fluorescein diacetate confirmation of viability (right) (Zhang et al 1998). Figure 6. Gallery of qRT-PCR of selected SSH-cDNA library members analyzed as in Fig. 5, with expression level of Sua set to 1. Clones were selected to represent differentially expressed gene products. A-L show some appreciable expression outside of sperm cells. F-M show upregulated expression in Sua. N-X show upregulated expression in Svn. *We acknowledge support by Bruce Roe, Sinkyoung So, Doris Kupfer, Tyrrell Conway and Mary Beth Langer in the OU AGCT Lab; Mohan Singh, Takashi Okada and Prem Bhalla at the University of Melbourne; and funding by USDA-NRI, Dept of Botany & Microbiology and Vice President for Research, University of Oklahoma. A