Download

1 / 48
480 likes | 1.07k Vues
Chémia sacharidov pre 5. ročník špecializácie Organická chémia, KOCH PRIF UK, Bratislava Sylaby kurzu.

E N D
Chémia sacharidovpre 5. ročník špecializácie Organická chémia, KOCH PRIF UK, Bratislava Sylaby kurzu 1. Výskyt sacharidov v prírode a ich rôznorodosť, chemické a biologické vlastnosti a význam, princípy glykobiológie, základné pojmy, definície a štruktúrne vyjadrenia, konformačná analýza, potreba špeciálnej nomenklatúry, vzťah k ostatnej organickej chémii. 2. Základné špeciálne reakcie sacharidov vyplyvajúce z ich polyhydroxykarbonylovej štruktúry. Mutarotácia, Lobry de Bruynova-Alberda van Ekensteinova reakcia, Bílikova reakcia. 3. Metódy skracovania a predlžovania uhlikového reťazca sacharidov. Fischerova glykozidácia. Vnútorné glykozidy (anhydridy sacharidov). Anomérny efekt.
Chémia sacharidovpre 5. ročník špecializácie Organická chémia Sylaby kurzu (pokračovanie) 4. Reakcie karbonylovej skupiny sacharidov s tiolmi, ditioacetály a ditioketály sacharidov, tioglykozidy. Reakcie karbonylovej skupiny sacharidov s amoniakom a jeho derivátmi. Amadoriho prešmyk, Maillardove reakcie. N-Glykozylamíny, hydrazóny a osazóny sacharidov. C-Glykozylované zlúčeniny. 5. Étery sacharidov. Metylétery, benzylétery, tritylétery, silylétery sacharidov, ich príprava, vlastnosti a využitie na protekciu a deprotekciu OH skupín sacharidov a na analytické účely. Metylačná analýza sacharidov. Jodistanová oxidácia a jej analytické využitie. NMR, hmotnostná spektrometria a chromatografické metódy analýzy sacharidov. 6. Estery sacharidov. Acetáty, benzoáty, pivaloáty, fosfáty, sulfáty, sulfonáty, boráty sacharidov, ich príprava, vlastnosti a využitie na protekciu a deprotekciu OH skupín sacharidov a na analytické účely.
Chémia sacharidovpre 5. ročník špecializácie Organická chémia Sylaby kurzu (pokračovanie 2) 7. Acetály sacharidov. Izopropylidénové a benzylidénové acetály, ich príprava, vlastnosti a využitie na protekciu a deprotekciu OH skupín sacharidov. Konformačná analýza izopropylidénových a benzylidénových acetálov. 8. Substitučné reakcie hydroxylových skupín sacharidov iných ako anomérnej OH skupiny. Využitie sulfonátov ako dobre odchadzajúcich skupín na túto substitúciu. Priama substitúcia hydroxylových skupín. 9. Anhydroderiváty sacharidov a ich príprava, vlastnosti a využitie na substitučné a konfiguračné manipulácie na uhlikovom skelete sacharidov. Pravidlá otvárania anhydrokruhov na rigidnych sacharidových štrukturách.
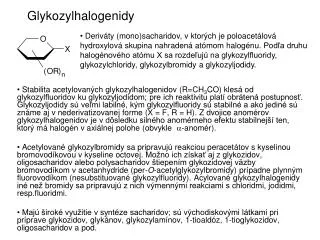
Chémia sacharidovpre 5. ročník špecializácie Organická chémia Sylaby kurzu (pokračovanie3) 10. Glykozylhalogenidy, glykozyltrichloracetimidáty, ich príprava, vlastnosti a využitie na syntézu glykozidov, disacharidov a vyšších oligosacharidov. 11. Tiosacharidy, aminodeoxysacharidy, neanomerické halogenidy sacharidov, ich príprava, vlastnosti a využitie. Sacharidy s rozvetveným reťazcom, nenasýtené deriváty sacharidov. 12. Využitie sacharidov ako zdrojov chirálnych syntónov na syntézu zlúčenín nesacharidovej povahy. Využitie sacharidov ako surovín chemického priemyslu v súčasnosti a vyhľad do budúcnosti. Nutričné vlastnosti sacharidov, stráviteľnosť a energetická výdatnosť sacharidov. Sladidlá a ich náhražky, náhražky tukov na báze sacharidov.
Literatúra:Monosaccharides. Their Chemistry and Their Rolesin Natural Products, P.M. Collins, R.J. Ferrier, Wiley, Chichester, 1995. Glycoscience I-III. Chemistry and Chemical Biology, B. Fraser-Reid, K. Tatsuta, J. Thiem (Eds.). Springer, Berlin, 2001. Contemporary Carbohydrate Chemistry, A. M. Stephen, University of Cape Town, RSA, 2003, http://web.uct.ac.za/depts/cem/ccc/contents.htm Sacharidy, M. Černý, T. Trnka, M. Budešínsky, Edice Chemických listů, ČSCH, 2010 Prednášky Chémia sacharidov pre 5. ročník špecializácie Organická chémia, KOCH PRIF UK, Bratislava http://chemia.rt.sk
Dva príklady zo záverečného testu 3.Konfigurácia D-gluko je • Medzistupňom chemických reakcií znázornených uvedenou reakčnou • schémou je štruktúra
Biologické funkcie sacharidov • Sacharidy,okremichuž dávno známych úloh v živej hmote, ktorými sú spomínané štrukturálno-stavebné a energeticko-zásobné funkcie, majú v živých organizmoch aj mnohé ďalšie, životne dôležité úlohy, ktoré veda začína spoznávať len v posledných desaťročiach. • Vďaka svojej štrukturálnej rozmanitosti, ktorá mnohonásobne prevyšuje štrukturálnu rozmanitosť nukleových kyselín a proteínov, sú sacharidy o.i. zodpovedné tak za rozpoznávacie procesy vo vnútri bunky ako i najejpovrchu, ale aj za špecializáciu buniek a s tým spojenú diferenciáciu tkanív a vývoj jednotlivých orgánov živých organizmov.
Schématická ilustrácia rozpoznávacích sacharidových interakcií medzi bunkami a medzi bunkou a vonkajšími činiteľmi. Zdroj: BioCarb AB katalóg.
Úloha povrchovobunkových oligosacharidov v rozpoznávacích prosesoch. (a) Bunky navzájom interagujú prostredníctvom viacnásobného viazania ich glykoproteínových oligosacharidov s proteínovými receptormi iných buniek. Glykolipidové (b) a glykoproteínové oligosacharidy (c) sú rozpoznávané protilátkami. Adhézia a kolonizácia bakteriálnych patogénov (d) na bunkách sa uskutočňuje prostredníctvom rozpoznania povrchových oligosacharidov. (e) Rovnako patogénne vírusy pred inváziou do buniek sa zachytávajú na povrchových oligosacharidoch špeciálnych pre daný vírus.
Schématické znázornenie bunkovej steny cicavčích buniek. Zdroj: http://cellbio.utmb.edu/cellbio/membrane_intro.htm resp. Wolfe S.L., Molecular and Cellular Biology, Wadsworth Publishing Company, 1993.)
Rôzne typy buniek spôsobujú produkciu rôznych súprav glykozylačných enzýmov, a zároveň majú rozličné sady proteínov, ktoré môžu byť glykozylované. • Glykozylácia je preto flexibilným evolučným nástrojom, ktorý slúži na široký rad účelov v zložitých organizmoch.
Najčastejšie monosacharidy cicavčích glykokonjugátov D-glukóza (D-Glc) D-galaktóza (D-Gal) D-manóza (D-Man) L-fukóza (L-Fuc) D-glukurónová kyselina (D-GlcA) D-xylóza (D-Xyl) L-idurónová kyselina (L-IdoA) N-acetyl-D-glukózamín (D-GlcNAc) N-acetyl-D-galaktózamín (D-GalNAc) N-acetyl-neuramínová(sialová) kyselina (Neu5Ac)
Monosacharidy cicavčích glykoproteínov. Prvých deväť štruktúr je najbežnejších, vyskytujú sa aj ďalšie, napr. NeuNGc a Kdn
Na rozdiel od cukrov sa základné stavebné jednotky nukleových kyselín a proteínov sa môžu viazať iba jedným typom chemickej väzby a môžu vytvárať iba nerozvetvené reťazce Nukleové kyseliny jediná kombinácia tymidín-mononukleotid Amino- kyseliny jediná kombinácia L-alanín + 7 iných kombinácií šesťčlenných kruhov (+ 31 iných kombinácií 5-a 6-členných kruhov) Cukry D-glukóza
Na rozdiel od cukrov sa základné stavebné jednotky nukleových kyselín a proteínov sa môžu viazať iba jedným typom chemickej väzby a môžu vytvárať iba nerozvetvené reťazce Nukleové kyseliny jediná kombinácia tymidín-mononukleotid Amino- kyseliny jediná kombinácia L-alanín + 7 iných kombinácií šesťčlenných kruhov (+ 31 iných kombinácií 5-a 6-členných kruhov)) Cukry D-glukóza
Cukry však vďaka svojej štrukturálnej a väzbovej rôznorodosti môžu vytvárať rozvetvené štruktúry • Takto relatívne jednoduchá sada monosacharidov môže vytvárať obrovský počet zložitých štruktúr • Napr. deväť bežných monosacharidov, ktoré sa nachádzajú v ľudskom organizme, môže byť poskladaných do viac ako 15 miliónov možných tetrasacharidov, z ktorých každý z nich môže byť považovaný za relatívne jednoduchý oligosacharid. • Tento glykokód sa využíva na rozpoznávacie procesy vo vnútri bunky i na jej povrchu, ale aj na špecializáciu buniek a s tým spojenú diferenciáciu tkanív, pletív a vývin jednotlivých orgánov živých organizmov.
Všetky bunky daného organizmu obsahujú rovnaký genetický materiál (genóm). • Od neho odvodený proteínový výkonný aparát (proteóm) ako aj sacharidové zloženie a funkcie (glykóm) sú však pre bunky špecifické podľa ich špecializácie a funkcie v organizme, čím sa bunky daného organizmu navzájom líšia. • Každá odchýlka od tohto normálneho sacharidového zloženia bunky potom spôsobuje chorobné zmeny v príslušnom tkanive, pletive či orgáne daného organizmu. Preto je poznanie sacharidového kódu jednotlivých buniek, teda diagnostika glykómu bunky, ako aj poznanie glykobiologických procesov v bunke, potrebné na kontrolu správnej činnosti a nevyhnutné na prípadnú potrebu opravy porúch proteínového aparátu bunky, ktorý na základe genetického kódu bunky zabezpečuje jej kompletnú výstavbu vrátane sacharidového zloženia.
Transfúzia krvi a objav krvných skupín Najznámejšia a medicínsky najdôležitejšia klasifikácia krvných skupín je podľa AB0 systému determinantov. AB0 krvné skupiny objavil Karl Landsteiner na Viedeňskej univerzite v rokoch 1900-1901, keď zisťoval príčinu, prečo transfúzia krvi niekedy spôsobí smrť a niekedy pacienta zase zachráni. V roku 1930 dostal za tento objav Nobelovu cenu. Karl Landsteiner(1868-1943)
Podľa tabuľky rozdelenia AB0 krvných skupín napr. ľudia s krvnou skupinou A majú na povrchu ich červených krviniek antigén A. Kvôli tomu neprodukujú anti-A protilátky, pretože tieto by spôsobili zničenie ich vlastnej krvi. Ak sa však do ich krvného obehu transfúziou zavedie krv krvnej skupiny B, anti-B protilátky prítomné v ich krvi (A) ju rozpoznajú ako cudziu a zničia ju alebo z nej aglutinujú (vyzrážajú do zhlukov) vnesené cudzie červené krvinky, a to za účelom očistenia krvi od cudzích proteínov.
Určenie krvnej skupiny:Neznáme ľudské červené krvinky pred pridaním (vľavo) a po pridaní (vpravo) séra obsahujúceho anti-A protilátky. Aglutinačná reakcia prezrádza prítomnosť antigénu A na povrchu týchto červených krviniek, čo dokazuje krvnú skupinu A (protilátky sú bielkoviny, červené krvinky sú bunky, ich špecifické antigény sú tetrasacharidy alebo pentasacharidy) http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/B/BloodGroups.html
Determinanty ľudských krvných skupín AB0 systému N-acetyl-D-galaktózamín L-fukóza červená krvinka Legenda: N-acetyl-D-glukózamín D-galaktóza
Sacharidový kód determinantovľudských krvných skupín AB0 systému rozpoznávajú bielkovinové protilátky na princípe klúča a zámku http://www.pandasthumb.org/archives/images/lock&key.jpg • Rovnaký princíp biologického rozpoznávania na bunkovej úrovni, najčastejšie medzi sacharidmi a bielkovinami, funguje aj v mnohých ďalších normálnych i chorobných procesoch v živých organizmoch
Všetky bunky daného organizmu obsahujú rovnaký genetický materiál (genóm). • Od neho odvodený proteínový výkonný aparát (proteóm) ako aj sacharidové zloženie a funkcie (glykóm) sú však pre bunky špecifické podľa ich špecializácie a funkcie v organizme, čím sa bunky daného organizmu navzájom líšia. • Každá odchýlka od tohto normálneho sacharidového zloženia bunky potom spôsobuje chorobné zmeny v príslušnom tkanive, pletive či orgáne daného organizmu. Preto je poznanie sacharidového kódu jednotlivých buniek, teda diagnostika glykómu bunky, ako aj poznanie glykobiologických procesov v bunke, potrebné na kontrolu správnej činnosti a nevyhnutné na prípadnú potrebu opravy porúch proteínového aparátu bunky, ktorý na základe genetického kódu bunky zabezpečuje jej kompletnú výstavbu vrátane sacharidového zloženia. • Rovnako dôležité je poznanie sacharidových kódov pri proteín-sacharidových interakciách buniek s exogénnymi patogénmi (baktérie, vírusy, toxíny), ktoré vstupujú do buniek práve na základe tohto biologického rozpoznania sacharidového kódu na svojom povrchu či na povrchu hostiteľských buniek. Na základe poznania týchto sacharidových kódov je potom možné úspešne navrhnúť lieky (glykoterapeutiká) na liečenie, ale aj na prevenciu ochorení, ktoré spôsobujú tieto patogény.
Kyselina N-acetylneuramínová (sialová) - prírodzený sacharid (cukor) nachádzajúci sa v cicavčích glykoproteínoch a glykolipidoch Zanamivír - aktívnazložka prvého protichrípkového lieku Relenza a účinný inhibítor enzýmu neuraminidáza, povrchového glykoproteínu chrípkových vírusov, ktorý štiepi koncové kyseliny sialové z glykoproteínov a glykolipidov a je kritický pre replikáciu vírusov Vírus chrípky pred inváziou do bunky
Druhým, dnes už do medicínskej praxe zavedeným protichrípkovým liekom, je prípravok Tamiflu, ktorého aktívnou zložkou je oseltamivír-fosfát, iný účinný inhibítor enzýmu neuraminidáza. Oseltamivír-fosfát je podobne ako predošlý prípravok Relenza (zanamivír) napodobneninou (mimetikom) kyseliny N-acetylneuramínovej. Oseltamivír-fosfát Kyselina N-acetylneuramínová (sialová)
Oseltamivir is an antiviral drug that slows the spread of influenza (flu) virus between cells in the body by stopping the virus from chemically cutting ties with its host cell—median time to symptom alleviation is reduced by 0.5–1 day. The drug is sold under the trade nameTamiflu and is taken orally in capsules or as a suspension. It has been used to treat and preventInfluenzavirus A and Influenzavirus B infection in over 50 million people since 1999. The prodrug Oseltamivir is itself not virally effective; however, once in the liver, it is converted by natural chemical processes, hydrolysed hepatically to its active metabolite, the free carboxylate of oseltamivir (GS4071). Oseltamivir is a neuraminidase inhibitor, serving as a competitive inhibitor towards sialic acid, found on the surface proteins of normal host cells. By blocking the activity of the viral neuraminidase enzyme, oseltamivir prevents new viral particles from being released by infected cells. http://en.wikipedia.org/wiki/Oseltamivir
Thiamet-G je veľmi sľubný kandidát na vývoj lieku proti Alzheimerovej chorobe Scott A Yuzwa, Matthew S Macauley, Julia E Heinonen, Xiaoyang Shan, Rebecca J Dennis, Yuan He, Garrett E Whitworth, Keith A Stubbs, Ernest J McEachern, Gideon J Davies & David J Vocadlo, Nature Chemical Biology 4, 483 - 490 (2008). Published online: 29 June 2008; doi:10.1038/nchembio.96 ( http://www.ksbi.or.kr/board/trendview.asp?BoardName=trend&Num=2278&ScrollAction=Page4
Thiamet G Hyperphosphorylation of the tau protein leads to its aggregation and formation of neurofibrillary tangles, a hallmark of Alzheimer’s disease and other tauopathic, neurodegenerative disorders. Tau is also dynamically modified by the addition and cleavage of O-linked β-N-acetylglucosamine (O-GlcNAc) moieties, which is mediated in part by O-GlcNAcase. Levels of O-GlcNAcylated proteins from Alzheimer’s disease brain extracts are decreased as compared to that in controls, suggesting that impaired brain glucose metabolism may contribute to pathogenesis. Thiamet G is a potent and selective inhibitor of O-GlcNAcase that demonstrates a Ki value of 21 nM. It increases cellular O-GlcNAc-modified protein levels (EC50 = 30 nM) and blocks phosphorylation of tau protein both in cultured PC-12 cells and in rats (200 mg/kg/day). Thiamet G is the first highly potent O-GlcNAcase inhibitor known to be orally bioavailable and effectively cross the blood brain barrier. http://www.caymanchem.com/app/template/Product.vm/catalog/13237/a/z;jsessionid=68481FD62E7985888DA013B37765339F
Monosacharidy cicavčích glykoproteínov. Prvých deväť štruktúr je najbežnejších, vyskytujú sa aj ďalšie, napr. NeuNAc a Kdn
Tri typy štruktúr asparagínovo-viazaných glykoproteínových oligosacharidov; 1 – komplexný, 2 – vysokomanózový, 3 – hybridný.Zdroj: http://www.prozyme.com/pdf/tngl100.pdf
Štyri typy základných štruktúr serínovo- resp. treonínovo-viazaných glykoproteínových oligosacharidov. Zdroj: http://www.prozyme.com/pdf/tngl100.pdf
Povrch bunky http://www.cdgs.com/glyco1.jpg
Základné pojmy, pôvod a význam sacharidov
sacharidy (syn. glycidy) - skupina zlúčenín s najmenej tromi alifaticky viazanými atómami uhlíka, ktorú tvoria polyhydroxykarbonylové zlúčeniny, ich jednoduché deriváty a ich oligoméry a polyméry, v ktorých monomérne sacharidové jednotky sú navzájom pospájané zmiešanou acetálovou (glykozidovou) väzbou. Podľa počtu jednoduchých sacharidových jednotiek obsiahnutých v jednej molekule sa sacharidy delia na monosacharidy, oligosacharidy a polysacharidy. K sacharidom sa zaraďujú aj glykokonjugáty (glykoproteíny, glykolipidy,atď.). Podľa prítomnosti karbonylovej skupiny v molekule sa sacharidy delia na redukujúce a neredukujúce. Sacharidy a ich deriváty obsahujujúce v molekule karbonylovú skupinu vo voľnej, hydratovanej alebo poloacetálovej forme sú redukujúce sacharidy. Sacharidy neobsahujúce takúto skupinu sú neredukujúce. Väčšina sacharidov sú bezfarebné (biele) kryštalické látky sladkej chuti. Rozpustnosť sacharidov je najlepšia vo vode a dimetylsulfoxide a klesá s klesajúcou polaritou rozpúšťadla. Reakcie sacharidov sa rozdeľujú na reakcie na hydroxylových skupinách, reakcie na karbonylovej skupine vo voľnej alebo poloacetálovej forme a reakcie podmienené prítomnosťou karbonylovej a hydroxylových skupín. Oligosacharidy a polysacharidy navyše poskytujú kyslou alebo enzýmovou hydrolýzou monosacharidy a oligosacharidy.
Na štruktúrnu analýzu sacharidov sa využívajú rôzne chemické reakcie, fyzikálnochemické metódy (1H- a 13C-NMR spektrometria, plynová a kvapalinová chromatografia, často v kombinácii s hmotnostnou spektrometriou, spektrometria v infračervenej, viditeľnej i ultrafialovej oblasti, polarimetria, optická rotačná disperzia, cirkulárny dichroizmus a rontgenová štruktúrna analýza) a biochemické metódy (reakcie katalyzované enzýmami). Sacharidy patria medzi najrozšírenejšie prírodné organické látky. Vznikajú v procese fotosyntézy (ročne asi 2 1011 ton), sú stavebným materiálom rastlín (50 až 80% sušiny) a základnou látkou metabolizmu živočíchov. Niektoré sacharidy sa získavajú z prírodných materiálov (D-glukóza, D-lyxóza, L-arabinóza, sacharóza, laktóza, atď.), ostatné sa z nich pripravujú synteticky. Využitie sacharidov je veľmi široké. Používajú sa ako potraviny a ich zložky (sacharóza, D-glukóza, D-fruktóza, laktóza, maltóza a maltodextríny, škrob, atď.), sú surovinou na výrobu textilných vlákien (celulóza), substrátmi pre rôzne fermentačné výroby (výroba etanolu, 1-butanolu, kyseliny citrónovej, atď.) a využívajú sa na výrobu liečiv, vitamínov a špeciálnych chemikálií. sacharidy (pokračovanie)
20 % 5 %, tuky, proteíny, terpenoidy, alkaloidy, atď.) lignín 75 % sacharidy Ročne v prírode fotosyntézou a následnými biochemickými premenami vzniká asi 200 miliárd ton biomasy, z ktorej prevažnú časť, asi 75 % tvoria sacharidy.
Historický vývoj základne chemického priemyslu _______________________________________________ Časové obdobie Surovinová základňa priemyselnej výroby materiálov_______________________________________________ Do prelomu Suroviny rastlinného pôvodu 19. a 20. storočia ↓ ↓ ↓ Obdobie od r. 1970 Suroviny fosílneho pôvodu ↓ ↓ ↓ ↓ ?? ?Suroviny rastlinného pôvodu?_______________________________________________
monosacharidy - zlúčeniny skupiny sacharidy, ktoré hydrolýzou neposkytujú nižšie sacharidové jednotky. Podľa typu karbonylovej skupiny sa monosacharidy delia na aldózy a ketózy. Ak je na sekundárny atóm uhlíka centrálneho uhlíkového reťazca viazaný ďalší uhlíkový atóm alebo uhlíkový reťazec, sú to monosacharidy s rozvetveným reťazcom. Základné derivaty monosacharidov sú deoxysacharidy, aminosacharidy, tiosacharidy, kyseliny aldónové, kyseliny urónové, kyseliny aldarové, alditoly a nenasýtené deriváty sacharidov (napr. glykály).K monosacharidom sa pričleňujú aj cyklitoly. Všetky tieto monosacharidy a ich deriváty môžu byť zložkami oligosacharidov a polysacharidov. Podľa konfigurácie na najvzdialenejšom asymetrickom atóme uhlíka (referenčný atóm uhlíka) od karbonylovej alebo potenciálnej karbonylovej skupiny sa rozdeľujú na D-monosacharidy a L-monosacharidy. Monosacharidy genetického radu D majú na referenčnom atóme uhlíka vo Fischerovom projekčnom vzorci hydroxylovú skupinu umiestnenú na pravej strane od centrálneho uhlíkového reťazca, monosacharidy genetického radu L majú túto hydroxylovú skupinu na ľavej srane. Reťazec uhlíkových atómov je orientovaný zvisle s uhlíkovým atómom karbonylovej alebo potenciálnej karbonylovej skupiny umiestneným hore. Väzby vystupujúce z atóma uhlíka k susedným atómom uhlíka v reťazci smerujú smerom za nákresnú rovinu, väzby k substituentom premietaným vpravo a vľavo od hlavného zvislého reťazca smerujú pred nákresnú rovinu. Atómy uhlíka v reťazci sa číslujú smerom zhora nadol.
monosacharidy - zlúčeniny skupiny sacharidy, ktoré hydrolýzou neposkytujú nižšie sacharidové jednotky. aldóza (D-ribóza) aldóza (D-glukóza) monosacharid s rozvetveným reťazcom (D-hamamelóza) ketóza (D-fruktóza) Redukujúce sacharidy vytvárajú cyklické poloacetálové formy.
monosacharidy - zlúčeniny skupiny sacharidy, ktoré hydrolýzou neposkytujú nižšie sacharidové jednotky. Podľa typu karbonylovej skupiny sa monosacharidy delia na aldózy a ketózy. Ak je na sekundárny atóm uhlíka centrálneho uhlíkového reťazca viazaný ďalší uhlíkový atóm alebo uhlíkový reťazec, sú to monosacharidy s rozvetveným reťazcom. Základné derivaty monosacharidov sú deoxysacharidy, aminosacharidy, tiosacharidy, kyseliny aldónové, kyseliny urónové, kyseliny aldarové, alditoly a nenasýtené deriváty sacharidov (napr. glykály).K monosacharidom sa pričleňujú aj cyklitoly. Všetky tieto monosacharidy a ich deriváty môžu byť zložkami oligosacharidov a polysacharidov. Podľa konfigurácie na najvzdialenejšom asymetrickom atóme uhlíka (referenčný atóm uhlíka) od karbonylovej alebo potenciálnej karbonylovej skupiny sa rozdeľujú na D-monosacharidy a L-monosacharidy. Monosacharidy genetického radu D majú na referenčnom atóme uhlíka vo Fischerovom projekčnom vzorci hydroxylovú skupinu umiestnenú na pravej strane od centrálneho uhlíkového reťazca, monosacharidy genetického radu L majú túto hydroxylovú skupinu na ľavej srane. Reťazec uhlíkových atómov je orientovaný zvisle s uhlíkovým atómom karbonylovej alebo potenciálnej karbonylovej skupiny umiestneným hore. Väzby vystupujúce z atóma uhlíka k susedným atómom uhlíka v reťazci smerujú smerom za nákresnú rovinu, väzby k substituentom premietaným vpravo a vľavo od hlavného zvislého reťazca smerujú pred nákresnú rovinu. Atómy uhlíka v reťazci sa číslujú smerom zhora nadol.
Základné deriváty monosacharidov: deoxysacharid(2-deoxy-D-ribóza) aminosacharid(N-acetyl-D-glukózamín) aldónová kyselina (k. D-glukónová) urónová kyselina(k. D-glukurónová) nenasýtený derivátsacharidu (D-glukál) cyklitol(myoinozitol) aldarová kyselina(k. D-glukarová k. cukrová) alditol (D-glucitol, sorbitol)
monosacharidy(pokračovanie) Monosacharidy a sacharidy vôbec ako chirálne molekuly (okrem mezo-foriem, ktoré majú rovinu symetrie, napr.galaktitol) sú opticky aktívne a otáčajú rovinu polarizovaného svetla. Optická otáčavosť [ ]D, znamená pri bežnej laboratórnej teplote (15 až 25o C) najčastejšie pri vlnovej dĺžke =589 nm (žltá D-čiara sodíkového svetla) vo vhodnom rozpúšťadle, je jednou z charakteristických konštánt pre tieto látky. Niekedy sa pravotočivosť (+) resp. ľavotočivosť (-) uvádza aj v názve sacharidu, napr. D(+)-glukóza. DL-Monosacharidy (napr. DL-glyceraldehyd) je ekvimolárna zmes obidvoch enanciomérov monosacharidov tzv. racemická zmes). Prítomnosť viacerých hydroxylových skupin v molekule monosachridu zvyšuje počet asymetrických centier v molekule a je príčinou existencie značného počtu stereoizomérov monosacharidov (enancioméry, diastereoizoméry, epiméry). Počet streoizomérov jednotlivých typov monosacharidov a ich derivátov pre štruktúru s n asymetrickými atómami uhlíka v acyklickej forme je daný pre nerovnako zakončený reťazec (napr. aldózy) čislom 2n (existuje napr. osem pentóz). Ak sú koncové skupiny v molekule rovnaké (napr. alditoly), je počet stereoizomérov daný číslom 2n-1 pre n nepárne (štyri pentitoly) resp. 2(n/2)-1. (2n/2+1) pre n párne (desať hexitolov). Redukujúce monosacharidy sú v acyklickej forme len v zriedkavých prípadoch. Ich charakteristickou vlastnosťou je vytváranie zmesí cyklických poloacetálových foriem (pyranóza, furanóza, ktoré sú vzájomne interkonvertovateľné (mutarotácia). Množstvo acyklickej formy v rovnováhe je spravidla veľmi malé. Novovytvorená hydroxylová skupina patriaca vnútornému poloacetálovému zoskupeniu sa nazýva anomérna (poloacetálová) hydroxylová skupina a sacharidový zvyšok glykozyl. Anomérny (poloacetálový) atóm uhlíka je ďalším asymetrickým centrom sacharidu.
In 1884 Fischer began his great work on the sugars, which transformed the knowledge of these compounds and welded the new knowledge obtained into a coherent whole. Even before 1880, the aldehyde formula of glucose had been indicated, but Fischer established it by a series of transformations such as oxidation into aldonic acid and the action of phenylhydrazine which he had discovered and which made possible the formation of the phenylhydrazones and the osazones. By passage to a common osazone, he established the relation between glucose, fructose and mannose, which he discovered in 1888. Hermann Emil Louis Fischer 1852-1919 (1902 Nobel Prize) In 1890, by epimerization between gluconic and mannonic acids, he established the stereochemical and isomeric nature of the sugars, and between 1891 and 1894 he established the stereochemical configuration of all the known sugars and exactly foretold the possible isomers, by an ingenious application of the theory of the asymmetrical carbon atom of Van't Hoff and Le Bel, published in 1874. Reciprocal syntheses between different hexoses by isomerization and then between pentoses, hexoses, and heptoses by reaction of degradation and synthesis proved the value of the systematics he had established. His greatest success was his synthesis of glucose, fructose and mannose in 1890, starting from glycerol. http://en.wikipedia.org/wiki/Hermann_Emil_Fischer
Organic and Medicinal Chemistry and Chirality • Practical significance since the times of Thalidomide (Contergan,1954-1957) http://en.wikipedia.org/wiki/Image:Thalidomide-structures.png
Vzťah medzi konformačným a Fischerovym vzorcom D-glyceraldehyd L-treóza
Optická aktivita sacharidov rovina symetrie galaktitol (meso-galactitol)[]D20 = 0 º D(+)-glukóza (dextróza)[]D20 = + 53 º D(-)-fruktóza(levulóza)[]D20 = – 92 º
Vzťah medzi počtom asymetrických uhlíkov a počtom izomérov sacharidov 8 aldopentóz 4 pentitoly Podobným spôsobom možno odvodiť súvislosť medzi 16 aldohexózami (8 D- a 8 L-enanciomérov) s 10 hexitolmi