Cell Death and Cell Renewal
830 likes | 1.79k Vues
17 Cell Death and Cell Renewal. Chapter OutlineProgrammed Cell DeathStem Cells and the Maintenance of Adult TissuesEmbryonic Stem Cells and Therapeutic Cloning. Introduction. Cell death and cell proliferation are balanced throughout the life of multicellular organisms.Animal development involves not only cell proliferation and differentiation but also cell death.Most cell death occurs by a normal physiological process of programmed cell death..

Cell Death and Cell Renewal
E N D
Presentation Transcript
1. Cell Death and Cell Renewal
2. 17 Cell Death and Cell Renewal Chapter Outline
Programmed Cell Death
Stem Cells and the Maintenance of Adult Tissues
Embryonic Stem Cells and Therapeutic Cloning
3. Introduction Cell death and cell proliferation are balanced throughout the life of multicellular organisms.
Animal development involves not only cell proliferation and differentiation but also cell death.
Most cell death occurs by a normal physiological process of programmed cell death.
4. Introduction In adult organisms, cell death must be balanced by cell renewal.
Most tissues contain stem cells that can replace cells that have been lost.
5. Programmed Cell Death Programmed cell death is carefully regulated.
In adults, it balances cell proliferation and maintains constant cell numbers.
It also eliminates damaged and potentially dangerous cells.
6. Programmed Cell Death During development, programmed cell death plays a key role by eliminating unwanted cells from a variety of tissues.
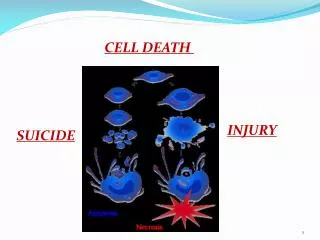
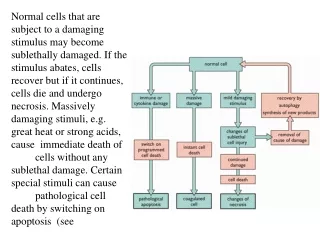
7. Programmed Cell Death Necrosis: Accidental cell death from acute injury.
Apoptosis: Programmed cell death; an active process.
Characterized by:
DNA fragmentation
Chromatin condensation
Fragmentation of the nucleus and cell
9. Programmed Cell Death Apoptotic cells and cell fragments are recognized and phagocytosed by macrophages and neighboring cells, and are rapidly removed from tissues.
Necrotic cells swell and lyse; the contents are released into the extracellular space and cause inflammation.
10. Programmed Cell Death Apoptotic cells express �eat me� signals, such as phosphatidylserine.
In normal cells, phosphatidylserine is restricted to the inner leaflet of the plasma membrane.
12. Programmed Cell Death Studies of C. elegans by the Robert Horvitz lab identified three genes with key roles in apoptosis.
C. elegans development includes the death of 131 specific cells.
Their experiments used mutant strains in which the cell death did not occur.
14. Programmed Cell Death The genes ced-3 and ced-4 were required for developmental cell death.
A third gene, ced-9, functioned as a negative regulator of apoptosis.
These genes are the central regulators and effectors of apoptosis that are highly conserved in evolution.
16. Programmed Cell Death Ced-3 is the prototype of a family of proteases known as caspases.
Caspases have cysteine (C) residues at their active sites and cleave after aspartic acid (Asp) residues in their substrate proteins.
17. Programmed Cell Death Caspases are the ultimate executioners of programmed cell death.
They bring about the events of apoptosis by cleaving 100 different cell target proteins.
The activation of an initiator caspase starts a chain reaction of caspase activation leading to death of the cell.
19. Programmed Cell Death Ced-4 and its mammalian homolog (Apaf-1) bind to caspases and promote their activation.
In mammalian cells, caspase-9 is activated by binding to Apaf-1 in a protein complex called the apoptosome.
Cytochrome c is also required, which is released from mitochondria.
21. Programmed Cell Death ced-9 in C. elegans is closely related to a mammalian gene called bcl-2, which was first identified as an oncogene.
Bcl-2 inhibits apoptosis. Cancer cells are unable to undergo apoptosis.
22. Programmed Cell Death Mammalian cells encode about 20 proteins related to Bcl-2, in three functional groups.
Some inhibit apoptosis, while others induce caspase activation.
The fate of the cell is determined by the balance of activity of proapoptotic and antiapoptotic Bcl-2 family members.
25. Programmed Cell Death In mammalian cells, members of the Bcl-2 family act at the mitochondria, which play a central role in controlling programmed cell death.
Cytochrome c is released from mitochondria, which triggers caspase activation in the apoptosome.
27. Programmed Cell Death Caspases are also regulated by a family of proteins called the IAP (inhibitor of apoptosis).
They either inhibit caspase activity or target caspases for ubiquitination and degradation in the proteasome.
29. Programmed Cell Death Regulation of programmed cell death is mediated by signaling pathways, some acting to induce cell death and others acting to promote cell survival.
Many forms of cell stress, such as DNA damage, can trigger programmed cell death.
30. Programmed Cell Death A major pathway leading to cell cycle arrest in response to DNA damage is mediated by the transcription factor p53.
Activation of p53 due to DNA damage can also lead to apoptosis.
32. Programmed Cell Death A major intracellular signaling pathway that promotes cell survival is initiated by the enzyme PI 3-kinase, which activates Akt.
Akt then phosphorylates a number of proteins that regulate apoptosis.
34. Programmed Cell Death Polypeptides in the tumor necrosis factor (TNF) family signal cell death by activating cell surface receptors.
These receptors directly activate a distinct initiator caspase, caspase-8.
37. Programmed Cell Death Programmed cell death can also occur by non-apoptotic mechanisms such as autophagy.
In normal cells, autophagy provides a mechanism for gradual turnover of the cell�s components by uptake of proteins or organelles into vesicles that fuse with lysosomes.
38. Programmed Cell Death Autophagy can also be an alternative to apoptosis as a pathway of cell death.
Autophagic cell death does not require caspases.
It can be activated by cellular stress and provide an alternative to apoptosis when apoptosis is blocked.
39. Programmed Cell Death Some forms of necrosis can be a programmed cellular response to stimuli such as infection or DNA damage.
Regulated necrosis may provide an alternative pathway of cell death if apoptosis does not occur.
40. Stem Cells and the Maintenance of Adult Tissues In early development, cells proliferate rapidly, then differentiate to form the specialized cells of adult tissues and organs.
To maintain a constant number of cells in adult tissues, cell death must be balanced by cell proliferation.
41. Stem Cells and the Maintenance of Adult Tissues Most differentiated cells in adult animals are no longer capable of proliferation.
If these cells are lost they are replaced by proliferation of cells derived from self-renewing stem cells.
42. Stem Cells and the Maintenance of Adult Tissues Some types of differentiated cells retain the ability to proliferate as needed, to repair damaged tissue throughout the life of the organism.
Fibroblasts in connective tissue can proliferate quickly in response to platelet-derived growth factor (PDGF) released at the site of a wound.
44. Stem Cells and the Maintenance of Adult Tissues Endothelial cells that line blood vessels can proliferate to form new blood vessels for repair and regrowth of damaged tissue.
46. Stem Cells and the Maintenance of Adult Tissues Endothelial cell proliferation is triggered by vascular endothelial growth factor (VEGF), which is produced by cells that lack oxygen.
48. Stem Cells and the Maintenance of Adult Tissues The epithelial cells of some internal organs are also able to proliferate to replace damaged tissue.
Liver cells, normally arrested in the G0 phase of the cell cycle, are stimulated to proliferate if large numbers of liver cells are lost (e.g., by surgical removal).
50. Stem Cells and the Maintenance of Adult Tissues Stem cells are less differentiated, self-renewing cells present in most adult tissues.
They retain the capacity to proliferate and replace differentiated cells throughout the lifetime of an animal.
51. Stem Cells and the Maintenance of Adult Tissues The key property of stem cells:
They divide to produce one daughter cell that remains a stem cell and one that divides and differentiates.
53. Stem Cells and the Maintenance of Adult Tissues Many types of cells have short life spans and must be continually replaced by proliferation of stem cells:
These include: blood cells, sperm, and epithelial cells of the skin and lining the digestive tract.
54. Stem Cells and the Maintenance of Adult Tissues Hematopoietic (blood-forming) stem cells were the first to be identified.
There are several distinct types of blood cells with specialized functions: erythrocytes, granulocytes, macrophages, platelets, and lymphocytes; all derived from the same population of stem cells.
56. Stem Cells and the Maintenance of Adult Tissues Epithelial cells that line the intestines live only a few days before they die by apoptosis.
New cells are derived from the continuous but slow division of stem cells at the bottom of intestinal crypts.
60. Stem Cells and the Maintenance of Adult Tissues Skin and hair are also renewed by stem cells.
The epidermis, hair follicles, and sebaceous glands are all maintained by their own stem cells.
62. Stem Cells and the Maintenance of Adult Tissues Stem cells also play a role in the repair of damaged tissue.
Skeletal muscle normally has little cell turnover, but it can regenerate rapidly in response to injury or exercise.
Regeneration is mediated by proliferation of satellite cells�the stem cells of adult muscle.
64. Stem Cells and the Maintenance of Adult Tissues Most adult tissues have stem cells, which reside in distinct microenvironments or niches.
Niches provide the environmental signals that maintain stem cells throughout life and control the balance between self-renewal and differentiation.
65. Stem Cells and the Maintenance of Adult Tissues Adult stem cells have potential utility in clinical medicine.
Hematopoietic stem cell transplantation (or bone marrow transplantation) plays an important role in the treatment of a variety of cancers.
67. Stem Cells and the Maintenance of Adult Tissues Epithelial stem cells are also used in the form of skin grafts to treat burns, wounds, and ulcers.
68. Embryonic Stem Cells and Therapeutic Cloning Embryonic stem cells can be grown indefinitely as pure stem cell populations that have pluripotency�the capacity to develop into all of the different types of cells in adult tissues.
Thus there is enormous interest in embryonic stem cells for both basic science and clinical applications.
69. Embryonic Stem Cells and Therapeutic Cloning Embryonic stem cells were first cultured from mouse embryos in 1981.
Mouse embryonic stem cells are an important experimental tool:
They can be used to introduce altered genes into mice.
They provide an outstanding model system for studying the molecular and cellular events associated with cell differentiation.
72. Embryonic Stem Cells and Therapeutic Cloning Human embryonic stem cell lines were first established in 1998.
Clinical transplantation therapies based on embryonic stem cells may provide the best hope for treatment of diseases such as Parkinson�s and Alzheimer�s disease, diabetes, and spinal cord injuries.
73. Embryonic Stem Cells and Therapeutic Cloning Mouse embryonic stem cells are grown in the presence of growth factor LIF, which is required to maintain the cells in their undifferentiated state.
If LIF is removed, the cells aggregate and differentiate.
Stem cells can be directed to differentiate along specific pathways by the addition of appropriate growth factors.
75. Embryonic Stem Cells and Therapeutic Cloning In 1997 Ian Wilmut and colleagues cloned Dolly the sheep.
Dolly arose by a process called somatic cell nuclear transfer.
This type of cloning in mammals is a difficult and inefficient process.
77. Embryonic Stem Cells and Therapeutic Cloning In therapeutic cloning, a nucleus from an adult human cell would be transferred to an enucleated egg.
The resulting embryo could produce differentiated cells for transplantation therapy.
This would bypass the problem of tissue rejection.
79. Embryonic Stem Cells and Therapeutic Cloning Problems to be overcome:
The low efficiency of generating embryos by somatic cell nuclear transfer.
Ethical concerns with respect to the possibility of cloning human beings (reproductive cloning), and with respect to the destruction of embryos.
80. Embryonic Stem Cells and Therapeutic Cloning These technical and ethical difficulties may be overcome by using induced pluripotent stem cells�reprogramming somatic cells to resemble embryonic stem cells.
The action of only four key transcription factors is sufficient to reprogram adult mouse somatic cells.
82. Embryonic Stem Cells and Therapeutic Cloning Adult human fibroblasts can be reprogrammed to pluripotency by a similar procedure.
Although problems remain, induced pluripotent stem cells may someday be used for patient-specific transplantation therapy.