Download
1 / 23
260 likes | 567 Vues
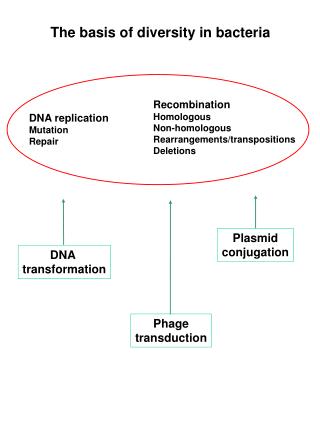
The basis of diversity in bacteria. Recombination Homologous Non-homologous Rearrangements/transpositions Deletions. DNA replication Mutation Repair. Plasmid conjugation. DNA transformation. Phage transduction. Transformation. Plating on selective medium (absence of Trp).

E N D
The basis of diversity in bacteria Recombination Homologous Non-homologous Rearrangements/transpositions Deletions DNA replication Mutation Repair Plasmid conjugation DNA transformation Phage transduction
Transformation Plating on selective medium (absence of Trp) DNA extracted from trp+ strain Transformed cells trp- strain Auxotrophy = inability of cells to grow unless the medium is supplemented with a particular substance Prototrophy = ability of cells to grow when a particular substance is absent from the medium The average fragment size of DNA is 32 Kb and transforming activity decreases with size The minimum required is 0,5 Kb. Synthesis of competence factor.
DNA associates with the membrane. Both Gram-positive and Gram-negative bacteria exhibit DNA binding, respectively. Double stranded breaks have been introduced into the entering DNA by the endogenous membrane-bound endonuclease. In addition DNA duplex is converted to single stranded DNA which recombines with chromosome.The region of mismatch is repaired by the DNA repair enzymes. In the example, lac- cells are transformed in lac+ cells.
In most system saturation is achieved by adding 1mg to 10 mg of DNA to 1 ml of culture. Transformation is a rare event in nature Competence, defined as the ability of a bacterium to take up DNA from the medium, can be naturally or artificially induced (divalent cation method for E. coli and related gram-negative bacteria, wall-degrading enzymes plus PEG for gram-positive bacteria, electroporation for eukaryotes and prokaryotes). Cells (N°/ml) Bacillus subtilis 2 Frequency of transformation (%) 1 0 time Natural competence B. subtilis, S. pneumoniae, H. influenzae, N. gonorrhoeae
Mediante trasformazione è possibile determinare la concatenazione di geni, la distanza e il loro ordine La probabilità di co-trasformazione è data dal prodotto delle probabilità di trasformazione dei singoli geni. Se due geni sono vicini è probabile trovarli sullo stesso frammento di DNA. La frequenza di co-trasformazione è inversamente proporzionale alla distanza tra i due geni. Quando 2 geni sono molto vicini, la loro frequenza di co-trasformazione è simile alla frequenza di trasformazione di ciascun gene Esempio: p+ q o = 2.5x10-3 p q+ o = 2.8x10-3 p q o+ = 2.6x10-3 p+ q o+ = 1.5x10-7 p+ q+ o = 1.5x10-3 p q+ o+ = 5x10-6 p q o • • •
Conjugation (the F factor) • Lederberg and Tatum demonstrated bacterial recombination (1946) This experiment was designed to determine whether mutation or recombination explained the appearance of new bacterial strains. • Davis’s U-tube experiments (1950) The pores in the filter were too small for bacteria to pass through but large enough to allow DNA molecules to pass freely. Lederberg experiment Davis experiment Conclusions: bacterial cell contact (coniugation) is required for DNA transfer and transformation could not explain recombination in E.coli. thr, treonina; bio, biotina; leu, leucina; thi, tiamina; met, metionina
Il trasferimento di DNA da un una cellula batterica ad un altra avviene in una sola direzione. F+ F- Hayes, nel 1953 propose l’esistenza di un fattore di fertilità (Fattore F). Il fattore F contiene i geni per la sintesi dei pili F o pili sessuali. Un ponte citoplasmatico mette in contatto la cellula donatrice con quella ricevente. La cellula F- è priva del fattore F. Coniugazione in E. coli Nell’ incrocio F+ x F- la cellula F- diventa F+. Cellula F- Cellula F+
1) An F+ cell may conjugate with an F- cell and transfer the F factor to the F- cell 2) Hfr (high frequency recombination) Luca Cavalli Sforza discovered Hfr strains The F plasmid is an episome 1) Recombination fr = 10-6 2) Recombination fr = 10-3
Time-of-entry gene mapping (interrupted mating) • Wollman & Jacob started the experiment with two E. coli strains (1956): • Hfr strain (donor) genotype • thr+ : Can synthesize threonine • leu+ : Can synthesize leucine • aziR : Resistant to azide • tonR : Resistant to the infection by T1 phage • lac+ : Can metabolize lactose • gal+ : Can metabolize galactose • strS : Killed by streptomycin • F– strain (recipient) genotype • thr–leu–aziS tonS lac–gal–strr thr, leu and str were genes allowing selection of recombinants while azi, ton, lac and gal were genes to be mapped
Recombinant cells were thr+, leu+ and strr. Recombinant colonies could be selected by culturing on medium containing streptomycin blocking the growth of Hfr strain and lacked threonine and leucine blocking the growth of F-strain. The researchers knew that the thr and leu genes were close to origin and that str gene was farther from the origin than the 4 genes to map.
Interpreting the Data Minutes that Bacterial Cells were Allowed to Mate Before Blender Treatment Percent of Surviving Bacterial Colonies with the Following Genotypes thr+ leu+ aziRtonR lac+ gal+ There were no surviving colonies after 5 minutes of mating 5 –– –– –– –– –– 10 100 12 3 0 0 15 100 70 31 0 0 20 100 88 71 12 0 25 100 92 80 28 0.6 30 100 90 75 36 5 40 100 90 75 38 20 50 100 91 78 42 27 60 100 91 78 42 27 1) After 10 minutes, the thr+ leu+genotype was obtained 3) It is followed by the tonR gene 2) The aziR gene is transferred first 4) The lac+ gene enters between 15 & 20 minutes 5) The gal+ gene enters between 20 & 25 minutes
The map of the four genes of interest (minutes) Complete transfer of the entire chromosomal DNA from Hfr cells requires 100 minutes in E. coli
The F-factor does not have a predetermined site on the chromosome for integration The F-factor orientation and map location of 25 Hfr strains of E. coli Detailed E. coli maps have been constructed using different F- and Hfr strains by means of time-of-entry mapping F- x Hfr
Arbitrarily assigned the starting point minutes Combining the results of mating experiments with different Hfr strains Campbell proposed that the chromosome of E. coli was circular.
The F plasmid is a large circular DNA, ~100 kb in length A large (~33 kb) region of the F plasmid, called the transfer region (~40 genes), is required for conjugation. 1) traJ is a positive regulator that turns on both traM and the traY-I operon. On the opposite strand, finP is a regulator that codes for a small antisense RNA that turns off traJ. 2) The genes traS and traT code for "surface exclusion" proteins. 3) The gene traA codes for the single subunit protein, pilin, that is polymerized into the pilus. 4) Transfer of the F factor is initiated at a site oriT while in its free form F utilizes its own replication origin oriV and is maintained at a level of one copy per bacterial chromosome. 5) There must be a channel through which DNA is transferred, but the pilus itself does not appear to provide it. TraD is an inner membrane protein in F+ bacteria that is necessary for transport of DNA and it may provide or be part of the channel.
F’ factors (sexduction) F’ factor is formed when different IS are used in excision and integration process. Once an F’ factor enters a recipient cell, the recipient cell has 2 copy of the piece of chromosomal DNA introduced on the F’ factor - merozygote or partial diploid. Complementation Analysis Experiments of Jacob and Monod using F’lac (1950-’60) Complementation tests are valuable in determining whether or not two mutations belong to the same gene.
Transduction Virulent phage enters the lytic cycle soon after infection. Temperate phage may enter the lytic cycle soon after infection or it may enter the lysogenic cycle, where it integrates (prophage) into the host cell chromosomal DNA and replicates as part of the chromosomal DNA. Plaques of T2 The discovery of transduction (Zinder and Lederberg, 1952) Salmonella typhimurium LA2 strain (phe+, trp+, met-, his-) Salmonella typhimurium LA22 strain (phe-, trp-, met+, his+) Presumably a substance passed across the filter and transmitted DNA (FA). FA (filterable agent) was not affected by DNase treatment neither transformation or conjugation could occur. A series of additional experiments, showed a consistent association between P22 and the observation of prototrophy, leading Zinder and Lederberg to conclude that the FA was the P22 phage. Salmonella LA22 strain was lysogenic for P22 phage while LA2 was lytic.
Specialized transduction phage particles contain mostly phage DNA but carry a specific fragment of chromosomal DNA () Generalized transduction phages particles contain DNA from any part of bacterial chromosome (P22, P1) Generalized Transduction
Specialized Transduction To enter the lysogenic condition, free lambda DNA must be integrated into the host DNA. To be released from lysogeny into the lytic cycle, prophage DNA must be excised from the chromosome. Integration occurs through the attachment sites. The sequence O (core) is common to attB and attP. The flanking regions B, B’ and P, P’ are referred to as the arms; each is distinct in sequence. The products of the recombination (prophage) are called attL and attR. attachment sites The reaction proceeds through a Holliday junction. Exchanges of single strands take place sequentially. The Integrase protein (Int), in the presence of IHF (Host Integration Factor), can resolve Holliday junctions, and is responsible for the cutting and ligation reactions.
Together, Int, Xis, and IHF cover virtually all of attP The function of attP requires a stretch of 240 bp, but the function of attB can be exercised by the 23 bp fragment extending from – 11 to +11, in which there are only 4 bp on either side of the core. The disparity in their sizes suggests that attP is providing additional information necessary to distinguish it from attB. 23 bp Int and IHF bind cooperatively to attP, and their affinity for the site is enhanced by supercoiling. The high stoichiometry suggests that the proteins do not function catalytically, but form a structure that supports only a single recombination event. When Int and IHF bind to attP, they generate a complex (Intasome) in which all the binding sites are pulled together on the surface of a protein. Supercoiling of attP is needed for the formation of this intasome. Int does not bind directly to attB in the form of free DNA. The intasome is the intermediate that "captures" attB. Int can form a similar complex with attR only if Xis is added. Such complex results in the excision process.
Formation of dbio and dgal transducing phages This event occurs at low frequency producing lambda defective phages Fr = 10-6 Fr = 10-6 Cell lysis LFT (Low Frequency of Transduction)
LFT is used to infect a lysogenic E.coli strain (gal-) In addition the gal or bio genes in phage may recombine with its homologous counterpart in the chromosome producing a gal+ or bio+ strains. HFT (High Frequency of Transduction by U.V. induction)