Download
1 / 28
310 likes | 565 Vues
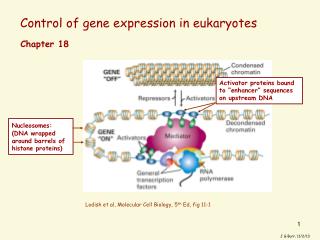
Control of gene expression in eukaryotes. Chapter 18. Activator proteins bound to “enhancer” sequences on upstream DNA. Nucleosomes: (DNA wrapped around barrels of histone proteins). Lodish et al, Molecular Cell Biology, 5 th Ed, fig 11-1. J G Burr, 11/2/13.

E N D
Control of gene expression in eukaryotes Chapter 18 Activator proteins bound to “enhancer” sequences on upstream DNA Nucleosomes:(DNA wrapped around barrels of histone proteins) Lodish et al, Molecular Cell Biology, 5th Ed, fig 11-1 J G Burr, 11/2/13
Remember we said that in the case of bacteria, the amount or activity of any protein found inside a bacterial cell could be governed in any of 3 ways: • transcriptional control • translational control • post-translational control J G Burr, 11/2/13
In the case of animal cells, there is also control at the level of: • Chromatin remodeling • Splicing of the primary RNA transcript • mRNA stability • A wider range of post-translational modification of proteins (eg, phosphorylation at serine, threonine and tyrosine sites; ubiquitination; glycosylation; acylation, prenylation, etc.) Once again, however, we are going to largely limit our focus to transcriptional regulation. J G Burr, 11/2/13
You may remember that bacterial promoters typically consist of a “Pribnow box” (-10 region) and a sequence called the “-35 region”: Bacterial promoter For most bacterial genes, the promoter is all that is necessary for RNA polymerase to sit down on the DNA and begin transcription. RNA Polymerase bound to the promoter. Fig 14.8, 1st Edition of your text J G Burr, 11/2/13
In other cases (eg, the lac operon) a DNA sequence that binds a positive control element like the CAP protein is required. For these genes, the promoter sequence is suboptimal, and binding of the CAP protein is necessary to recruit RNA polymerase. Transcription of eukaryotic genes is in general terms more like this. Eukaryotic RNA polymerase does not bind to its promoter without the assistance of other proteins, a little bit like the way that transcription of the lac operon requires CAP. J G Burr, 11/2/13
Eukaryotic promoters have a sequence that sort of resembles the “Pribnow box” of prokaryotic promoters, but it is located -25 to -35 to the transcription start site. It is called a “TATA box”. RNA polymerase cannot directly bind to the promoter (the TATA box); instead a protein called “TATA box-binding protein” (TBP) binds at this site. TBP induces a sharp bend in the DNA (just like the CAP protein in the case of bacteria). TBP is a subunit of a protein complex called “TFIID”. (TFIID = TBP + 13 “TAF” proteins.) (TAF = “TBP-Associated Factors”.) Lodish et al, Molecular Cell Biology, 5th Ed, Fig 11-27 J G Burr, 11/2/13
After TFIID binds to the TATA box and bends the DNA, a set of additional proteins (TFIIA, TFIIB,TFIIE, TFIIH) are recruited to the promoter. TFIID, TFIIA, TFIIB,TFIIE, & TFIIH are called “general transcription factors”, because they assemble at all promoters whenever mRNA is going to be transcribed. Once the general transcription factors have assembled on the promoter, RNA polymerase is able to bind. Alberts et al, Molecular Biology of the Cell, 4th Ed, Fig 6-16 J G Burr, 11/2/13
But the assembly of the general transcription factors on the promoter requires the assistance of additional gene regulatory proteins, called transcriptional activators. These proteins bind at any of several sites, sometimes adjacent to the promoter (“promoter-proximal elements”); sometimes at more distant locations. The binding sites at distant locations are called “enhancers”. Enhancers Promoter-proximal elements Enhancer Promoter Transcriptional activator proteins bind at these sites Lodish et al, Molecular Cell Biology, 5th Ed, Fig 11-12 J G Burr, 11/2/13
Here’s a representation of the binding of transcriptional activator proteins to enhancer sequences on the DNA, both upstream and downstream of the promoter. But how can these activator proteins assist in the assembly of general transcription factors on the promoter, since they are so far away? J G Burr, 11/2/13
Answer: the flexibility of DNA.The DNA between the enhancer and the promoter loops out to permit the activator proteins bound to an enhancer to assist in the assembly of the general transcription factors on the promoter: (Enhancer sequence) General Transcription Factors DNA J G Burr, 11/2/13
Most often the interaction between the activator proteins and the general transcription factors involves a protein complex called a “mediator”: J G Burr, 11/2/13
In addition to the activators and mediators that are required for eukaryotic gene expression, nucleosome structure must also be modified. Chromatin-remodeling proteins and histone acetylases come into play here. J G Burr, 11/2/13
Remember, eukaryotic DNA wraps around ‘barrels” of histone proteins to form “nucleosomes”. The combination of DNA and histones is called “chromatin”, and chromatin can exist both in an extended “beads on a string” state, and in more condensed forms, especially a condensed form called the “30nm fiber”. The 30nm fiber is the basic state of most chromatin in most cells. Genes are inaccessible for transcription in a 30nm fiber; they are accessible in the “beads-on-a-string” form. “Linker DNA” Nucleosomes J G Burr, 11/2/13
Chromatin-remodeling proteins and histone acetylases act to convert chromatin in the 30nm form to the more accessible “beads-on-a-string” form, which permits the binding of transcriptional activators, and permits RNA polymerase to navigate through nucleosomes. J G Burr, 11/2/13
Both histone acetylases and chromatin remodeling complexes act to locally convert 30nm chromatin fibers to beads-on-a-string chromatin. J G Burr, 11/2/13
In addition to unpacking 30nm fibers, chromatin remodeling complexes (a) reposition nucleosomes on the DNA [thereby often exposing enhancer sites] and (b) also loosen the DNA on the nucleosomes, permitting RNA polymerase to navigate through the nucleosomes: J G Burr, 11/2/13
Other chromatin remodeling complexes return the nucleosomes back to their standard state when gene expression is finished: J G Burr, 11/2/13
A little more detail on histones and nucleosomes: The nucleosome has a core of 8 histone proteins: 2 each of histones H2A, H2B, H3 and H4. J G Burr, 11/2/13
Each of the 4 types of histone protein that form the octameric core of a nucleosome has its amino terminus “waving free”. These “free-waving” amino terminal segments are rich in the positively charged amino acid lysine; and some of the lysines in these tails may play a role in the stabilization of 30nm fibers: 30nm fiber J G Burr, 11/2/13
It is the lysines in the amino-terminal histone segments that can become acetylated by the histone acetylases that play such an important role in chromatin remodeling and gene activation Acetylated lysine side chains are no longer positively charged; they would therefore no longer interact with DNA or other proteins in the same way. “K” = lysine Acetylated lysines can however now interact with new, different proteins. J G Burr, 11/2/13
Acetylation of these lysines has several functions in gene activation, including unpacking of the 30nm fiber, and also recruiting general transcription factors such as TFIID, once the 30nm fibers have become unpacked Unpacking Recruit RNA polymerase Lysines 8 & 16 on histone H4 have become acetylated J G Burr, 11/2/13
Broadly speaking, acetylation of histone lysines by histone acetyl transferases (HATs) is associated with decondensing chromatin and making genes accessible for transcription. And removing these acetyl groups from the histones, by histone deacetylases (HDACs), silences gene expression by repacking the nucleosomes back into 30nm fibers. Histone acetyl transferases activate genes Histone deacetylases silence genes J G Burr, 11/2/13
Here’s how your text summarizes transcriptional regulation in eukaryotes: J G Burr, 11/2/13
Before we move on to our next topic (cancer), we will briefly consider two other aspects of the regulation gene expression in eukaryotes: (1) Alternative splicing; and (2) RNA interference (micro RNAs). Lets look first at (1) alternative RNA splicing, using as an example the expression of two different versions of the protein tropomyosin in skeletal muscle versus smooth muscle. The two different proteins are encoded by a single gene, but the 1º RNA transcript is spliced differently in the two different cell types, to produce 2 mRNAs: Poly A tail Poly A tail Two different mRNAs encode two different (but related) proteins J G Burr, 11/2/13
Now let’s take a look at the role of a class of RNA molecules called micro RNAs (miRNA) in the regulation of gene expression. These miRNA molecules act to target certain mRNA molecules (to which they are complementary in sequence) for rapid degradation. J G Burr, 11/2/13
There are several hundred genes that encode miRNA molecules, that have evolved to target specific mRNAs in the cytosol, thereby limiting the lifetime of these mRNAs and curtailing expression of the protein encoded by the target mRNA.These miRNAs initially form a hairpin structure which is trimmed in the nucleus, then exported to the cytosol, where the loop at the end is cleaved, yielding ultimately a 22 nucleotide long double-stranded mature miRNA molecule. J G Burr, 11/2/13
In the cytosol, a set of proteins, the RISC complex, bind the double-stranded miRNA, removing one of the strands. The remaining strand is complementary to a sequence on the target mRNA. The miRNA-RISC complex then binds the target mRNA, and a ribonuclease component of the RISC complex cleaves the mRNA. If there is an imperfect match between the miRNA and the mRNA, the mRNA is not cleaved, but the RISC-miRNA complex remains bound to the mRNA, thereby inhibiting its translation. J G Burr, 11/2/13
And that’s the end of the material from your text that I’ll be lecturing on. We are now going to finish the course with a set of lectures on the molecular basis of cancer, and I’ll make my lecture notes on this materialavailable to you as usual as downloadable pdf files on our course home page and in the Bookstore Copy Center. Cancerous tumors Metastatic cancer in the liver J G Burr, 11/2/13