Download
1 / 29
320 likes | 454 Vues
Explore the dynamics of stomatal opening and closing, the role of potassium in turgor changes, and plant responses to external stimuli through tropisms and hormonal signaling pathways.

E N D
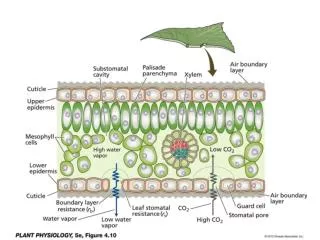
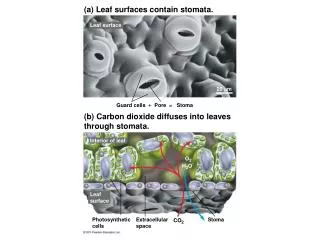
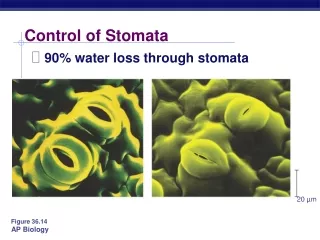
20 µm Figure 36.14 Control of Stomata • 90% water loss through stomata
Cells turgid/Stoma open Cells flaccid/Stoma closed (a) Changes in guard cell shape and stomatal opening and closing (surface view). Guard cells of a typical angiosperm are illustrated in their turgid (stoma open) and flaccid (stoma closed) states. The pair of guard cells buckle outward when turgid. Cellulose microfibrils in the walls resist stretching and compression in the direction parallel to the microfibrils. Thus, the radial orientation of the microfibrils causes the cells to increase in length more than width when turgor increases. The two guard cells are attached at their tips, so the increase in length causes buckling. Radially oriented cellulose microfibrils Cell wall Vacuole Guard cell Figure 36.15a • Guard cells change shape
H2O H2O H2O H2O (b) Role of potassium in stomatal opening and closing. The transport of K+ (potassium ions, symbolized here as red dots) across the plasma membrane and vacuolar membrane causes the turgor changes of guard cells. H2O K+ H2O H2O H2O H2O H2O Figure 36.15b • Changes in turgor pressure • Proton pumps =uptake of K+ = then H2O diffuses in = turgid = open
4 cues • Env stress to close • blue-green light receptor stimulates pumps to open • Depletion of CO2 in leaf space to open • Circadian rhythm
Chapter 39 • PLANT RESPONSES TO EXTERNAL AND INTERNAL STIMULI
bending of a grass seedling toward light • plant sensing the direction, quantity, and color of the light Figure 39.1
TROPISM • growth response = movement due to env. stimulus • Phototropism • Gravitropism • Roots = + gravitropism • Stems = - gravitropism • Thigmotropism
Statoliths 20 m (a) (b) gravitropism • detect gravity by the settling of statoliths • Specialized plastids containing dense starch grains Figure 39.25a, b
thigmotropism • Rubbing the stems of young plants a couple of times daily • Results in plants that are shorter than controls Figure 39.26
How do plants “know” to respond to all these things? • Tropisms and other plant activity = due to hormones: • coordinate cell division, • elongation, • differentiation • responses to stimuli
CYTOPLASM CELL WALL 3. Response 1. Reception 2. Transduction Activation of cellular responses Relay molecules Receptor Hormone or environmental stimulus Plasma membrane Figure 39.3 CELL SIGNALING • response to stimulus = signals cell processing Trigger Gene Expression To increase Protein synthesis Proteins Relay “message” = cascade Protein changes
SIGNAL TRANSDUCTION PATHWAY Receptors = sensitive to very weak environmental OR chemical signals secondary messengers - internally produced chemicals • transfer and amplify the signal from receptor to other proteins • EX: cGMP (Fig. 39.4)
SIGNAL TRANSDUCTION PATHWAY Light signal is detected by phytochrome receptor which activates 2 signal transduction pathways Ca++ and cGMP = 2nd messengers that activate proteinkinases Both pathways turn on genes that produce proteins thatproduce response
EXPERIMENT In 1880, Charles Darwin and his son Francis designed an experiment to determine what part of the coleoptile senses light. In 1913, Peter Boysen-Jensen conducted an experiment to determine how the signal for phototropism is transmitted. Boysen-Jensen (1913) Control Darwin and Darwin (1880) Shaded side of coleoptile Light RESULTS Light Light Base covered by opaqueshield Tip separated by gelatinblock Tip separated by mica Illuminated side of coleoptile Tip removed Tip covered by opaque cap Tip covered by trans-parentcap CONCLUSION In the Darwins’ experiment, a phototropic response occurred only when light could reach the tip of coleoptile. Therefore, they concluded that only the tip senses light. Boysen-Jensen observed that a phototropic response occurred if the tip was separated by a permeable barrier (gelatin)but not if separated by an impermeable solid barrier (a mineral called mica). These results suggested that the signal is a light-activated mobile chemical. phototropism Figure 39.5
EXPERIMENT In 1926, Frits Went’s experiment identified how a growth-promoting chemical causes a coleoptile to grow toward light. He placed coleoptiles in the dark and removed their tips, putting some tips on agar blocks that he predicted would absorb the chemical. On a control coleoptile, he placed a block that lacked the chemical. On others,he placed blocks containing the chemical, either centered on top of the coleoptile to distribute the chemical evenly or offset to increase the concentration on one side. RESULTS The coleoptile grew straight if the chemical was distributed evenly. If the chemical was distributed unevenly, the coleoptile curved away from the side with the block, as if growing toward light, even though it was grown in the dark. Excised tip placed on agar block Growth-promotingchemical diffusesinto agar block Agar blockwith chemicalstimulates growth Control(agar blocklackingchemical)has noeffect Offset blockscause curvature Control CONCLUSION Went concluded that a coleoptile curved toward light because its dark side had a higher concentration of the growth-promoting chemical, which he named auxin. • Extracted the chemical messenger for phototropism, auxin, by modifying earlier experiments • In 1926, Frits Went Figure 39.6
1. Auxin • promotes cell elongation • Most abundant in meristimatic tissues
control of apical dominance • terminal bud suppresses development of axillary buds Figure 39.9a
2. Gibberellins • over 100 ID’d • Effects • stem elongation • fruit growth • seed germination plump grapes in grocery stores have been treated with gibberellin hormones while on the vine
4. Abscisic Acid • Promote seed dormancy • Drought tolerance
One bad apple spoils the whole bunch… 5. Ethylene • In response to stresses: • drought, flooding, mechanical pressure, injury, and infection • Promotes aging and dropping of leaves in Fall • Ripens fruit
Apoptosis in plants • response to hormones • ethylene • auxin • death of annual plant after flowering • differentiation of xylem vessels • loss of cytoplasm • shedding of autumn leaves
Photoperiodism • Light cues key events in plant activity • Circadian rhythm • response to length of day/night • 2 classes of Proteins that respond to light…..
1.0 0.8 0.6 Phototropic effectiveness relative to 436 nm 0.4 0.2 0 450 500 550 600 650 700 400 Wavelength (nm) Light Time = 0 min. Time = 90 min. 1. Blue-light photoreceptors Induce phototropism, or opening stomata Researchers exposed maize (Zea mays) coleoptiles to violet, blue, green, yellow, orange, and red light to test which wavelengths stimulate the phototropic bending toward light.
A phytochrome consists of two identical proteins joined to form one functional molecule. Each of these proteins has two domains. Chromophore Photoreceptor activity. One domain, which functions as the photoreceptor, is covalently bonded to a nonprotein pigment, or chromophore. Kinase activity. The other domain has protein kinase activity. The photoreceptor domains interact with the kinase domains to link light reception to cellular responses triggered by the kinase. Figure 39.19 2. phytochromes Proteins that absorb mostly red light Light triggers change in structure!!! = triggers kinase activity of other molecules
2 forms of phytochrome • Conversion between isomer forms • Pr = exposure to red light (660 nm) • Inactive form • Accumulates at night • no germination • Pfr = far-red (730 nm) • Active form • Accumulates during day • germination
Pr Pfr Red light Responses: seed germination, control of flowering, etc. Synthesis Far-red light Slow conversion in darkness (some plants) Enzymatic destruction
DAY LENGTH & FLOWERING • SHORT DAY PLANTS • need daylight for less than a “critical period” to flower • EX: poinsettia • (flower in late summer, fall, winter) • LONG DAY PLANTS • -need daylight for longer thana certain “critical period” • EX: lettuce • (flower in late spring/early summer)
NIGHT LENGTH = CRITICAL “Short day plants” (=really need LONG NIGHT) don’t flower if dark time is interrupted by short burst of light
Flowering hormone mystery!!! • Florigen??? • Not yet identified!!! • Hormone produced in leaves • Thought to be transported from leaves to apical meristem to initiate flowering