Why is gene expression tightly regulated?
580 likes | 824 Vues
What are the differences between prokaryotes and eukaryotes on the initiation of TRANSCRIPTION and TRANSLATION?. Why is gene expression tightly regulated?. For prokaryotes, it is the way that organisms adapt to the environment. For eukaryotes

Why is gene expression tightly regulated?
E N D
Presentation Transcript
What are the differences between prokaryotes and eukaryotes on the initiation of TRANSCRIPTION and TRANSLATION?
Why is gene expression tightly regulated? • For prokaryotes, it is the way that organisms adapt to the environment. • For eukaryotes • Gene expression differs among cell types: cell differentiation • Is involved in development • Is associated with physiology and pathology • Genotype+expression level (affected by environment)=phenotype
Gene expression regulation is different between prokaryotes and eukaryotes due to the differences in • Cellular structure • Existing form of genomic DNA • Complexity in gene expression
Regulation of gene expression Organization of bacterial genes The lac operon Catabolite repression The trp operon Attenuation Regulation by alternative sigma factors
1. Regulation of gene expression Regulation of gene expression allows bacteria to respond to changes in their environment, typically to the presence or absence of nutrients. Balance between the rate of synthesis [‘sinθisis] and the rate of degradation [ degrə'deɪʃən]
Factors that alter the rate of synthesis: Changes in the rate of gene transcription, the best understood Changes in mRNA turnover time(更新的时间) Changes in the rate of translation
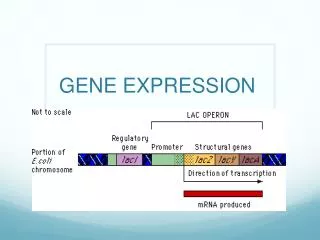
2. Organization of bacterial genes Important feature: operon['ɔpə,rɔn], transcriptional units in which several genes, usually encoding proteins with related functions, are regulated together 操纵子:是原核生物基因表达和调控的单位,由启动子、操纵基因(调控)及其所控制的一组功能上相关的结构基因所组成。操纵基因受调节基因产物的控制。
In 1961, Jacob and Monod found the difference between two types of sequences in DNA: sequences that code for trans-acting products(反式作用因子); and cis-acting sequences(顺式作用序列) that function exclusively within the DNA. Gene activity is regulated by the specific interactions of the trans-acting products (usually proteins) with the cis-acting sequences (usually sites in DNA).
A gene (反式作用 products) a sequence of DNA that codes for a product. This product may be protein (as in the case of the majority of genes) or may be RNA (as in the case of genes that code for tRNA and rRNA etc.). • The crucial feature is that the product • diffuses away from its site of synthesis to • act elsewhere. 一种基因的蛋白质产物,能影响位于基因组另一条染色体上的(或基因组别处的)另一个基因的活性,这种作用叫做反式作用。参与这种反式作用的特定因子叫做反式作用因子,一般指的是蛋白质,它能够通过与顺式作用元件(DNA)之间的相互作用而影响基因的表达。
The cis-acting sequence functions exclusively as a DNA sequence, affecting only the DNA to which it is physically linked. (In some cases, a cis-acting sequence functions in an RNA rather than in a DNA molecule, such as RNA termintor) 影响自身基因表达活性的非编码DNA序列。
Classification of operons: Inducible operons(可诱导操纵子): contain genes that encode enzymes involved in metabolic pathways. e.g. lac operon Repressible operons(可阻遏操纵子): contain genes that encode enzymes involved in biosynthetic pathways and gene expression is controlled by the end product of the pathway e.g. trp operon
Positive regulation(正调控):有调节蛋白(激活蛋白)存在时基因表达开启,没有调节蛋白存在时则基因的表达关闭; Negative regulation(负调控):在调节蛋白(阻遏蛋白)存在时基因的表达关闭,无调节蛋白时基因的表达得以开启。 正、负调控中调节蛋白都是反式作用因子,通常识别基因上有的顺式作用元件
3. The lac operon Three enzymes for the metabolism of lactose: Lactose permeaseβ半乳糖苷通透酶(Lac Y): transport lactose into the cell β-galatosidaseβ半乳糖苷酶(Lac Z): hydrolyzes lactose into glucose and galactose (把乳糖分解为葡萄糖和半乳糖) Transacetylaseβ半乳糖苷转乙酰基酶(Lac A): hydrolysis[hai’drɔlisis]
Z编码β-半乳糖苷酶:将乳糖水解成葡萄糖和半乳糖Z编码β-半乳糖苷酶:将乳糖水解成葡萄糖和半乳糖 • Y编码β-半乳糖苷透过酶:使外界的β-半乳糖苷(如乳糖)能透过大肠杆菌细胞壁和原生质膜进入细胞内。 • A编码β-半乳糖苷乙酰基转移酶:乙酰辅酶A上的乙酰基转到β-半乳糖苷上,形成乙酰半乳糖。
Regulatory Gene Operon DNA I P O Z Y A m-RNA Protein β-Galactosidase Transacetylase Permease Control elements -5 +21 repressor
Lac Z, Y and Aare transcribed as a single mRNA (多顺反子mRNA) from a single promoter. Lac Ilies upstream of the operon and encodes lac repressor, which regulates the expression of Lac Z, Y and A genes. LacI 表达阻遏物。操纵基因(O)是DNA上的一小段序列(仅为26bp),是阻遏物的结合位点。
Regulatory Gene Operon DNA I P O Z Y A m-RNA Protein β-Galactosidase Transacetylase Permease Control elements +21 -5 repressor
Lac repressor -5 +21 Plac Lac repressor is encoded by LacI, which isactive as atetramerof identical subunits. It occupies the operator-binding site Olac (28bp, palindromic) and blocks almostly all transcription of lacZYA when lack of inducer (such as lactose).
In the absence of lactose Lac repressor binds to operator Blocks the path of the RNA polymerase move through the lac promoter and operator Prevents the transcription of lac genes
When lactose present • A few molecules of the lac enzymes present in the cell allow lactose to be taken up and metabolized • Lactose becomes allolactose, the latter binds to the lac repressor and changes its conformation such that it can no longer binds to the operator • The block is removed and the operon is transcribed • When the lactose is used up, lac repressor returns to its original conformation, again block transcription
4. Catabolite repression Catabolite repression: the presence of glucose in the cell switches off the lac operon 葡糖糖存在时,只利用其作为能量代谢来源物质,对乳糖的存在视而不见,因而乳糖相关的酶类也无需表达。 葡萄糖可以抑制腺苷酸环化酶的活性。 In the presence of cAMP, CAP (catabolite activator protein) binds to a DNA sequence upstream of the lac promoter and enhances binding of the RNA polymerase leading to the enhanced transcription of the operon 当细菌处于碳饥饿状态时(葡萄糖缺乏时),腺苷酸环化酶转化ATP为cAMP, cAMP 的量显著提高,可以与其激活蛋白CAP(或者叫CRP)形成二聚体,并与特定的DNA序列结合起始操作子进行转录。形成正调控体系
Hey man, I’m constitutive Repressor Promoter LacZ LacY LacA CAP Binding Repressor Repressor Repressor mRNA Operator CAP The Lac Operon:When Glucose Is Present But Not Lactose Come on, let me through RNA Pol. No way Jose!
Hey man, I’m constitutive RNA Pol. Repressor Promoter LacZ LacY LacA X CAP CAP CAP Binding Repressor Repressor Repressor mRNA cAMP cAMP cAMP Repressor Operator CAP The Lac Operon:When Lactose Is Present But Not Glucose Bind to me Polymerase Yipee…! RNA Pol. Lac This lactose has bent me out of shape
Alright, I’m off to the races . . . Hey man, I’m constitutive Repressor Promoter LacZ LacY LacA CAP CAP CAP Binding Repressor Repressor Repressor mRNA cAMP cAMP cAMP Operator CAP The Lac Operon:When Neither Lactose Nor Glucose Is Present Bind to me Polymerase Come on, let me through! RNA Pol. STOP Right there Polymerase
乳糖操纵子的正调控(catabolite repression) • When glucose absent • Adenylate cyclase is active and cAMP levels are higher • CAP binds, lac operon transcription level increases
When glucose present • Adenylatecyclase腺苷酸环化酶) is inhibited and cAMP levels are low。 • CAP does not bind, transcription of the lac operon is very low
Summary A C B Glucose/lactose Repressor/operator binding cAMP-CAP/promoter A: RNA polymerase B: lac repressor C: CRP-cAMP
5. The trp operon Structure five genes encoding enzymes involved in biosynthesis of tryptophan Leader Trp operator Trp promoter Trp repressor lies upstream of the trp operon 当环境中存在可利用的色氨酸时,细菌会减少和停止自身的合成,通过Trp 操纵子来实现这种阻遏。
3.4.1.2 Tryptophan Metabolism in E. coli: A Repressible Gene System trpE、D、C、B和A分别编码: 邻氨基苯甲酸合成酶的ε和δ亚基、吲哚甘油磷酸合成酶、色氨酸合成酶的β和α链。它们都是合成色氨酸必需的原料。
When tryptophan present Tryptophan binds to trp repressor enabling it to binds to trp operator Obstruct binding of RNA polymerase to the trp promoter Prevent transcription (变构)
When tryptophan absent • Trp repressor is incapable of binding to trp operator, transcription proceeds
6. Attenuation(衰减作用) Expression level is finely tuned.(与翻译过程相偶联的转录调控模型)
Structure of attenuator(衰减子的结构) • Trp mRNA: 14 codons followed by a stop codon, two consecutivetrp codons • Trp L: including four complementary sequences which can form two stem-loop structures. The small one terminates transcription (ρ-factor independent terminator), while the large one not.
抗终止信号 终止信号
If levels of tryptophan are adequate • Trp mRNA is translated, ribosome prevents formation of larger stem-loop, transcription ends
If tryptophan is lacking • The ribosome will be stalled as it translates the coding region • RNA polymerase will move ahead, form larger stem-loop, transcription can proceed
Low Trp High Trp
Attenuation determines how efficiency it is transcribed. Attenuation depends on the fact that transcription and translation are linked in bacteria.
7. Regulation by alternative sigma factors Sigma factor (σ) is responsible for initiating transcription by recognizing bacterial promoter DNA sequences. By using different sigma factors, different sets of genes are transcribed to meet different environmental conditions.
factor subunit bound to RNA pol for transcription initiation Released core enzyme 2’ RNA elongation factors: is bifunctional protein • Bind to core RNA Pol • 识别启动子共有序列并且只为转录起始所需 • (-35 and –10) in DNA
Promoter recognition 多种不同的σ factors binding to the same RNA Pol 细胞中RNA聚合酶可以识别不同的启动子,形成不同的转录物 70factors is most common one in E. coli under the normalgrowth condition