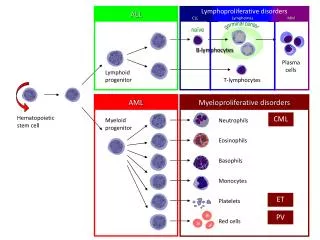
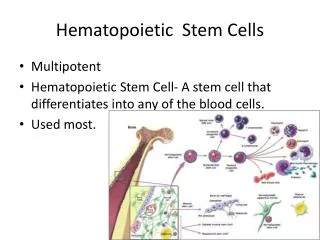
Hematopoietic Stem Cell Mobilization
E N D
Presentation Transcript
1. Hematopoietic Stem Cell Mobilization Armin Ghobadi, MD
Fellow, Hematology & Oncology
Washington University in St Louis
10/28/2011 1
2. Disclosure
I have NO financial relationships to
disclose 2
3. Overview History
Stem cell niche
Pathways/receptors involving stem cell mobilizaion and homing
GCSF
GCSF + Chemotherapy
CXCR-4 Inhibition
Plerixafor (Mozobil)
Other agents
VLA-4 inhibition
Summary
References
3
4. History 1957: Thomas et al. IV infusion of bone marrow in patients receiving radiation and
chemotherapy ( N. Engl. J. Med. 257, 491�496 (1957))
1958: Bond et al. DNA synthesizing cells in peripheral blood of normal human beings
(Science 1958;128:202-203)
1961: Bierman et al. Leukapheresis in Men (Brit. J. haematol., 7:51-63, 1961)
1962: Goodman et al. Evidence for stem cells in the peripheral blood of mice
(Blood 1962;19:702-714)
1971: McCredie et al. Cells capable of colony formation in the peripheral blood of man
(Science 1971;171:293-4)
1979/1980: Hershko et al and Abrams et al. Failed PBSCT attempts
(Lancet 1979;1(8123):945-947 and Blood 1980;56:516-520)
1981: Goldman et al. First successful autologous PBSCT (N Engl J Med 1981;305:700)
1985: Nicola et al. Human G-CSF was introduced (Nature 314:625, 1985)
1988: D�hrsen et al. G-CSF increases PB progenitor cells up to 100 fold
(Blood 1988;72(6):2074-2081)
1989: Kessinger et al: First allogeneic PBSCT. (Bone Marrow Transplant.1989;4(6):643-646.)
1991: Filgrastim was approved for chemotherapy induced neuropenia
1992: Sheriden et al. PBSCT using G-CSF as a mobilizing agent
2008/2009: DiPersio et al. 2 randomized phase 3 trials confirmed AMD3100 (Plerixafor) safety and efficacy
in NHL and MM (Blood 2009;113(23):5720-5726 and J Clin Oncol 2009;27(28):4767-4773)
AMD3100 (Plerixafor) was FDA-approved for HSPC mobilization in combination with G-CSF 4 the term �blood stem cell� as a transplantable cell population was introduced in 1962
The Nobel Prize in Physiology or Medicine 1990 E. Donnall Thomas
1971: McCredie et al: first attempt at M. D. Anderson Cancer Center of collecting colony-forming stem cells by continuous-flow apheresis was successful
Micklem HS, Anderson N, Ross E. Limited potential of circulating haemopoietic stem cells. Nature 1975;256:41-3.
---1979/1980: Two clinical blood stem-cell transplantation attempts among identical twins, one in 1979 at the University of California at Los Angeles12 and the other in 1980 at the NIH,13 resulted in failed or incomplete lymphohematopoietic reconstitution. In the University of California at Los Angeles case the leukapheresis product was administered to the twin patient as 6 separate white blood cell transfusions during the course of 14 days. The total amount of mononuclear cells transfused was 5.8 � 1010/kg recipient body weight. More than 2 months after transfusion, no platelet or granulocyte engraftment was documented. One may speculate that each individual white blood cell transfusion did not contain a sufficient amount of hematopoietic progenitor cells to enable engraftment. In the NIH case, the leukapheresis-derived mononuclear cells were administered as daily infusions during the course of 8 days. No recovery of granulocytes, monocytes, or platelets was documented at 8 weeks after transfusion. The authors attributed engraftment failure to the low number of clonogenic hematopoietic progenitor cells transfused being only one-eighth the quantity used in BM transplantation. Subsequently, both patients were rescued with their twin's marrow cells. It should be noted that, at that time, stem-cell collection by apheresis was performed at steady state without any growth factor priming
1981: in CML using leukopheresis and cryopreservationthe term �blood stem cell� as a transplantable cell population was introduced in 1962
The Nobel Prize in Physiology or Medicine 1990 E. Donnall Thomas
1971: McCredie et al: first attempt at M. D. Anderson Cancer Center of collecting colony-forming stem cells by continuous-flow apheresis was successful
Micklem HS, Anderson N, Ross E. Limited potential of circulating haemopoietic stem cells. Nature 1975;256:41-3.
---1979/1980: Two clinical blood stem-cell transplantation attempts among identical twins, one in 1979 at the University of California at Los Angeles12 and the other in 1980 at the NIH,13 resulted in failed or incomplete lymphohematopoietic reconstitution. In the University of California at Los Angeles case the leukapheresis product was administered to the twin patient as 6 separate white blood cell transfusions during the course of 14 days. The total amount of mononuclear cells transfused was 5.8 � 1010/kg recipient body weight. More than 2 months after transfusion, no platelet or granulocyte engraftment was documented. One may speculate that each individual white blood cell transfusion did not contain a sufficient amount of hematopoietic progenitor cells to enable engraftment. In the NIH case, the leukapheresis-derived mononuclear cells were administered as daily infusions during the course of 8 days. No recovery of granulocytes, monocytes, or platelets was documented at 8 weeks after transfusion. The authors attributed engraftment failure to the low number of clonogenic hematopoietic progenitor cells transfused being only one-eighth the quantity used in BM transplantation. Subsequently, both patients were rescued with their twin's marrow cells. It should be noted that, at that time, stem-cell collection by apheresis was performed at steady state without any growth factor priming
1981: in CML using leukopheresis and cryopreservation
5. Allo-HSCT source - Adults 5
6. Allo-HSCT Source - Pediatrics 6
7. Auto-HSCT source 7
8. HSC Niche
Sinusoidal endothelial cells
1. VCAM-1
2. CXCL12 8 --Maintenance of HSCs and
regulation of their self-renewal and differentiation in vivo
is thought to depend on their specific microenvironment,
which has been historically called the haematopoieticinductive
microenvironment10 or �stem-cell niche�
--Models of HSPC mobilization by G-CSF and inhibitors of CXCR4 and VLA-4. (a) Bone marrow environment at baseline. HSPCs in the
endosteal niche are likely in close contact with osteoblasts and nestin? MSCs, both of which express numerous HSPC retention factors, including
CXCL12, VCAM-1 and SCF. The perivascular niche is more distant from the endosteum and includes both nestin? MSCs and CXCL12 abundant
reticular cells. Cells of the monocytic lineage support the maintenance of osteoblasts and MSCs. b-Adrenergic nerve cells of the sympathetic
nervous system regulate MSC proliferation and induce circadian oscillations of CXCL12 expression. (b) Model of G-CSF-induced HSPC
mobilization. Cells of the monocytic lineage express the receptor for G-CSF and provide factors that support the survival of MSCs and osteoblasts.
Upon 4�5 days of stimulation with G-CSF, the monocytes/macrophages disappear, leading to the loss of osteoblast lineage cells and reduced
expression of CXCL12, VCAM-1 and SCF on MSCs. Reduced expression of these important HSPC retention factors is also observed following
G-CSF signaling through b-adrenergic nerve cells. The net effect of these signaling cascades is the disruption of HSPC retention interactions and
mobilization of HSPCs into the peripheral blood. (c, d) Model of HSPC mobilization by inhibitors of CXCR4 or VLA-4. Targeted disruption of the
interaction of CXCR4 or VLA-4 with their ligands results in the rapid (within hours) and reversible mobilization of HSPCs into the peripheral
circulation. An additive or synergistic affect on HSPC mobilization is observed when a CXCR4 inhibitor is combined with a VLA-4 antagonist,
G-CSF or their combination. When used alone, inhibitors of CXCR4 and VLA-4 mobilize fewer HSPCs than G-CSF.
-----------------------------------------------------
Support for the hypothesis that one cell type is present at various sites within the marrow niche was obtained by a recent landmark study by M�ndez-Ferrer et al. (2010). This group identified a stromal nestin-expressing MSC population (nestin+ MSC) that is closely associated with putative HSCs. Nestin+ MSCs are strictly perivascular and are typically found in more central areas of the marrow, but they are also present in the immediate vicinity of the endosteum, albeit at lower frequency. Moreover, nestin+ MSCs are tightly associated with adrenergic nerve fibers of the sympathetic nervous system (SNS) that regulate HSC mobilization and are responsible for the circadian oscillations in circulating HSC numbers (Katayama et al., 2006; M�ndez-Ferrer et al., 2008). Strikingly, these MSCs express higher levels of HSC maintenance factor transcripts, including CXCL12, stem cell factor (SCF), angiopoietin-1 (Ang-1), IL-7, vascular cell adhesion molecule 1 (VCAM1), and osteopontin (OPN), compared with any other stromal cell type including OBs. Depletion of nestin+ MSCs by inducible expression of diphtheria toxin receptor (DTR) in nestin-expressing cells caused the mobilization of ~50% of HSCs to the spleen. In addition, the homing of transplanted progenitor cells into MSC-depleted recipients was reduced by 90%.Moreover, treatment with the HSC-mobilizing factor granulocyte colony-stimulating factor (G-CSF) decreased the expression of CXCL12 and other factors required for HSC maintenance and retention by nestin+ MSCs. These cells also seem to mediate the capacity of parathyroid hormone to increase HSC numbers, which had previously been correlated with an increase in putative niche OBs (Calvi et al., 2003; Adams et al., 2007). Parathyroid hormone directly stimulated proliferation of nestin+ MSCs while simultaneously promoting their differentiation into OBs (M�ndez-Ferrer et al., 2010). Collectively, these data support the hypothesis that nestin+ MSCs are a functional component of the bone marrow HSC niche.Most interestingly, nestin+ MSCs show several similarities to recently identified mesenchymal progenitors (Sugiyama et al., 2006; Omatsu et al., 2010). These bipotent adipoosteogenic progenitors were identified in a mouse strain in which GFP was expressed from the endogenous CXCL12 locus (Sugiyama et al., 2006; Omatsu et al., 2010). Because of their high CXCL12 expression and their long cellular processes, these cells were named CXCL12-abundant reticular (CAR) cells. The majority of putative HSCs are found in close proximity to CAR cells by immunohistochemistry, and like nestin+ MSCs, CAR cells are predominantly found in the more central areas of the marrow, with some also located close to vessels near the endosteum (Fig. 1). Although CAR cells are more abundant than nestin+ MSCs, they too are tightly associated with sinusoidal endothelium and have a similar morphology to vascular pericytes. These data are in agreement with studies in humans, suggesting that virtually all MSC activity is found within the larger pericyte population that associates closely with the vascular system in the entire body (Crisan et al., 2008).Similar to nestin+ MSCs, CAR cells express HSC maintenance proteins such as CXCL12 and SCF. Induced depletion of CAR cells using a DTR approach causes a partial loss of HSC activity associated with a decrease in HSC cycling, suggesting that CAR cells promote HSC cycling and self-renewal. Even though this phenomenon was not observed after depletion of nestin+ MSCs, it seems likely that CAR cells and nestin+ MSCs represent two highly overlapping CXCL12-expressing cell populations. Because nestin+ MSCs are approximately four times less abundant than CAR cells, contain all colony-forming-unit fibroblast activity within the marrow, harbor high self-renewal activity in vitro and in vivo, and are capable of multilineage differentiation into bone, cartilage, and fat, it appears that nestin+ MSCs may represent a more primitive population compared with CAR cells and may even be a CAR subpopulation. As it remains unclear whether nestin+ MSCs are homogeneous and whether they all express high levels of CXCL12, it is possible that some nestin+ MSCs may not be CAR cells. Although future studies will need to dissect the cellular relationship between these two niche cell populations, there is now convincing evidence that perivascular mesenchymal stem/progenitor cells are critical inhabitants of the HSC niche. (Edinger et al. JEM vol. 208 no. 3 421-428 )
----SNS promotes egress by reducing the expression of key retention factors by the niche cell (M�ndez-Ferrer et al., 2010b), and in contrast, BM Macrophage promotes the expression of these genes and HSC/progenitor retention in the BM.--Maintenance of HSCs and
regulation of their self-renewal and differentiation in vivo
is thought to depend on their specific microenvironment,
which has been historically called the haematopoieticinductive
microenvironment10 or �stem-cell niche�
--Models of HSPC mobilization by G-CSF and inhibitors of CXCR4 and VLA-4. (a) Bone marrow environment at baseline. HSPCs in the
endosteal niche are likely in close contact with osteoblasts and nestin? MSCs, both of which express numerous HSPC retention factors, including
CXCL12, VCAM-1 and SCF. The perivascular niche is more distant from the endosteum and includes both nestin? MSCs and CXCL12 abundant
reticular cells. Cells of the monocytic lineage support the maintenance of osteoblasts and MSCs. b-Adrenergic nerve cells of the sympathetic
nervous system regulate MSC proliferation and induce circadian oscillations of CXCL12 expression. (b) Model of G-CSF-induced HSPC
mobilization. Cells of the monocytic lineage express the receptor for G-CSF and provide factors that support the survival of MSCs and osteoblasts.
Upon 4�5 days of stimulation with G-CSF, the monocytes/macrophages disappear, leading to the loss of osteoblast lineage cells and reduced
expression of CXCL12, VCAM-1 and SCF on MSCs. Reduced expression of these important HSPC retention factors is also observed following
G-CSF signaling through b-adrenergic nerve cells. The net effect of these signaling cascades is the disruption of HSPC retention interactions and
mobilization of HSPCs into the peripheral blood. (c, d) Model of HSPC mobilization by inhibitors of CXCR4 or VLA-4. Targeted disruption of the
interaction of CXCR4 or VLA-4 with their ligands results in the rapid (within hours) and reversible mobilization of HSPCs into the peripheral
circulation. An additive or synergistic affect on HSPC mobilization is observed when a CXCR4 inhibitor is combined with a VLA-4 antagonist,
G-CSF or their combination. When used alone, inhibitors of CXCR4 and VLA-4 mobilize fewer HSPCs than G-CSF.
-----------------------------------------------------
Support for the hypothesis that one cell type is present at various sites within the marrow niche was obtained by a recent landmark study by M�ndez-Ferrer et al. (2010). This group identified a stromal nestin-expressing MSC population (nestin+ MSC) that is closely associated with putative HSCs. Nestin+ MSCs are strictly perivascular and are typically found in more central areas of the marrow, but they are also present in the immediate vicinity of the endosteum, albeit at lower frequency. Moreover, nestin+ MSCs are tightly associated with adrenergic nerve fibers of the sympathetic nervous system (SNS) that regulate HSC mobilization and are responsible for the circadian oscillations in circulating HSC numbers (Katayama et al., 2006; M�ndez-Ferrer et al., 2008). Strikingly, these MSCs express higher levels of HSC maintenance factor transcripts, including CXCL12, stem cell factor (SCF), angiopoietin-1 (Ang-1), IL-7, vascular cell adhesion molecule 1 (VCAM1), and osteopontin (OPN), compared with any other stromal cell type including OBs. Depletion of nestin+ MSCs by inducible expression of diphtheria toxin receptor (DTR) in nestin-expressing cells caused the mobilization of ~50% of HSCs to the spleen. In addition, the homing of transplanted progenitor cells into MSC-depleted recipients was reduced by 90%.Moreover, treatment with the HSC-mobilizing factor granulocyte colony-stimulating factor (G-CSF) decreased the expression of CXCL12 and other factors required for HSC maintenance and retention by nestin+ MSCs. These cells also seem to mediate the capacity of parathyroid hormone to increase HSC numbers, which had previously been correlated with an increase in putative niche OBs (Calvi et al., 2003; Adams et al., 2007). Parathyroid hormone directly stimulated proliferation of nestin+ MSCs while simultaneously promoting their differentiation into OBs (M�ndez-Ferrer et al., 2010). Collectively, these data support the hypothesis that nestin+ MSCs are a functional component of the bone marrow HSC niche.Most interestingly, nestin+ MSCs show several similarities to recently identified mesenchymal progenitors (Sugiyama et al., 2006; Omatsu et al., 2010). These bipotent adipoosteogenic progenitors were identified in a mouse strain in which GFP was expressed from the endogenous CXCL12 locus (Sugiyama et al., 2006; Omatsu et al., 2010). Because of their high CXCL12 expression and their long cellular processes, these cells were named CXCL12-abundant reticular (CAR) cells. The majority of putative HSCs are found in close proximity to CAR cells by immunohistochemistry, and like nestin+ MSCs, CAR cells are predominantly found in the more central areas of the marrow, with some also located close to vessels near the endosteum (Fig. 1). Although CAR cells are more abundant than nestin+ MSCs, they too are tightly associated with sinusoidal endothelium and have a similar morphology to vascular pericytes. These data are in agreement with studies in humans, suggesting that virtually all MSC activity is found within the larger pericyte population that associates closely with the vascular system in the entire body (Crisan et al., 2008).Similar to nestin+ MSCs, CAR cells express HSC maintenance proteins such as CXCL12 and SCF. Induced depletion of CAR cells using a DTR approach causes a partial loss of HSC activity associated with a decrease in HSC cycling, suggesting that CAR cells promote HSC cycling and self-renewal. Even though this phenomenon was not observed after depletion of nestin+ MSCs, it seems likely that CAR cells and nestin+ MSCs represent two highly overlapping CXCL12-expressing cell populations. Because nestin+ MSCs are approximately four times less abundant than CAR cells, contain all colony-forming-unit fibroblast activity within the marrow, harbor high self-renewal activity in vitro and in vivo, and are capable of multilineage differentiation into bone, cartilage, and fat, it appears that nestin+ MSCs may represent a more primitive population compared with CAR cells and may even be a CAR subpopulation. As it remains unclear whether nestin+ MSCs are homogeneous and whether they all express high levels of CXCL12, it is possible that some nestin+ MSCs may not be CAR cells. Although future studies will need to dissect the cellular relationship between these two niche cell populations, there is now convincing evidence that perivascular mesenchymal stem/progenitor cells are critical inhabitants of the HSC niche. (Edinger et al. JEM vol. 208 no. 3 421-428 )
----SNS promotes egress by reducing the expression of key retention factors by the niche cell (M�ndez-Ferrer et al., 2010b), and in contrast, BM Macrophage promotes the expression of these genes and HSC/progenitor retention in the BM.
9. Endosteal Niche-Stem Cell Synapse 9 A model of the endosteal niche�stem-cell synapse. Schematic diagram of the endosteal niche�stem-cell
synapse showing putative ligand�receptor interactions and adhesion molecules, as well as some of the intracellular
pathways that are activated following signalling. ANG1, angiopoietin-1; BMI1, polycomb repressor; BMP, bone
morphogenetic protein; BMPR1A, BMP receptor 1A; CSL, CBF1 suppressor of Hairless and LAG1; CXCL12, CXCchemokine
ligand 12; CXCR4, CXC-chemokine receptor 4; FAK, focal adhesion kinase; HOXB4, homeobox B4; HSC,
haematopoietic stem cell; ICAM1, intercellular adhesion molecule 1; LFA1, lymphocyte function-associated antigen 1;
LRP, low-density-lipoprotein-receptor-related protein; MAPK, mitogen-activated protein kinase; OPN, osteopontin;
PI3K, phosphatidylinositol-3 kinase; PLC, phospholipase C; PKC, protein kinase C; PPR, PTH/PTH-related protein receptor;
PTH, parathyroid hormone; SCF, stem-cell factor; SMADS, mothers against decapentaplegic-related homologue;
SNO, spindle-shaped N-cadherin-expressing osteoblast; TIE2, tyrosine kinase receptor 2; VCAM1, vascular cell-adhesion
molecule 1; VLA4, very late antigen 4; �?� denotes molecules and/or interactions for which only indirect or contradictory
evidence is available.A model of the endosteal niche�stem-cell synapse. Schematic diagram of the endosteal niche�stem-cell
synapse showing putative ligand�receptor interactions and adhesion molecules, as well as some of the intracellular
pathways that are activated following signalling. ANG1, angiopoietin-1; BMI1, polycomb repressor; BMP, bone
morphogenetic protein; BMPR1A, BMP receptor 1A; CSL, CBF1 suppressor of Hairless and LAG1; CXCL12, CXCchemokine
ligand 12; CXCR4, CXC-chemokine receptor 4; FAK, focal adhesion kinase; HOXB4, homeobox B4; HSC,
haematopoietic stem cell; ICAM1, intercellular adhesion molecule 1; LFA1, lymphocyte function-associated antigen 1;
LRP, low-density-lipoprotein-receptor-related protein; MAPK, mitogen-activated protein kinase; OPN, osteopontin;
PI3K, phosphatidylinositol-3 kinase; PLC, phospholipase C; PKC, protein kinase C; PPR, PTH/PTH-related protein receptor;
PTH, parathyroid hormone; SCF, stem-cell factor; SMADS, mothers against decapentaplegic-related homologue;
SNO, spindle-shaped N-cadherin-expressing osteoblast; TIE2, tyrosine kinase receptor 2; VCAM1, vascular cell-adhesion
molecule 1; VLA4, very late antigen 4; �?� denotes molecules and/or interactions for which only indirect or contradictory
evidence is available.
10. G-CSF/Filgrastim-model of HSPC mobilization 10 Models of HSPC mobilization by G-CSF and inhibitors of CXCR4 and VLA-4. (a) Bone marrow environment at baseline. HSPCs in the
endosteal niche are likely in close contact with osteoblasts and nestin? MSCs, both of which express numerous HSPC retention factors, including
CXCL12, VCAM-1 and SCF. The perivascular niche is more distant from the endosteum and includes both nestin? MSCs and CXCL12 abundant
reticular cells. Cells of the monocytic lineage support the maintenance of osteoblasts and MSCs. b-Adrenergic nerve cells of the sympathetic
nervous system regulate MSC proliferation and induce circadian oscillations of CXCL12 expression. (b) Model of G-CSF-induced HSPC
mobilization. Cells of the monocytic lineage express the receptor for G-CSF and provide factors that support the survival of MSCs and osteoblasts.
Upon 4�5 days of stimulation with G-CSF, the monocytes/macrophages disappear, leading to the loss of osteoblast lineage cells and reduced
expression of CXCL12, VCAM-1 and SCF on MSCs. Reduced expression of these important HSPC retention factors is also observed following
G-CSF signaling through b-adrenergic nerve cells. The net effect of these signaling cascades is the disruption of HSPC retention interactions and
mobilization of HSPCs into the peripheral blood. (c, d) Model of HSPC mobilization by inhibitors of CXCR4 or VLA-4. Targeted disruption of the
interaction of CXCR4 or VLA-4 with their ligands results in the rapid (within hours) and reversible mobilization of HSPCs into the peripheral
circulation. An additive or synergistic affect on HSPC mobilization is observed when a CXCR4 inhibitor is combined with a VLA-4 antagonist,
G-CSF or their combination. When used alone, inhibitors of CXCR4 and VLA-4 mobilize fewer HSPCs than G-CSF.
-- Models of HSPC mobilization by G-CSF and inhibitors of CXCR4 and VLA-4. (a) Bone marrow environment at baseline. HSPCs in the
endosteal niche are likely in close contact with osteoblasts and nestin? MSCs, both of which express numerous HSPC retention factors, including
CXCL12, VCAM-1 and SCF. The perivascular niche is more distant from the endosteum and includes both nestin? MSCs and CXCL12 abundant
reticular cells. Cells of the monocytic lineage support the maintenance of osteoblasts and MSCs. b-Adrenergic nerve cells of the sympathetic
nervous system regulate MSC proliferation and induce circadian oscillations of CXCL12 expression. (b) Model of G-CSF-induced HSPC
mobilization. Cells of the monocytic lineage express the receptor for G-CSF and provide factors that support the survival of MSCs and osteoblasts.
Upon 4�5 days of stimulation with G-CSF, the monocytes/macrophages disappear, leading to the loss of osteoblast lineage cells and reduced
expression of CXCL12, VCAM-1 and SCF on MSCs. Reduced expression of these important HSPC retention factors is also observed following
G-CSF signaling through b-adrenergic nerve cells. The net effect of these signaling cascades is the disruption of HSPC retention interactions and
mobilization of HSPCs into the peripheral blood. (c, d) Model of HSPC mobilization by inhibitors of CXCR4 or VLA-4. Targeted disruption of the
interaction of CXCR4 or VLA-4 with their ligands results in the rapid (within hours) and reversible mobilization of HSPCs into the peripheral
circulation. An additive or synergistic affect on HSPC mobilization is observed when a CXCR4 inhibitor is combined with a VLA-4 antagonist,
G-CSF or their combination. When used alone, inhibitors of CXCR4 and VLA-4 mobilize fewer HSPCs than G-CSF.
--
11. The most common mobilization strategies Allo-HSCT:
G-CSF
Auto-HSCT:
G-CSG
GM-CSF
Chemomobilization (Cytoxan + G-CSF)
Pleraxifor + G-CSF for NHL and MM
11
12. G-CSF 10 mcg/kg day or 5 mcg/kg BID
Start leukaphaeresis on day 5
Minimum number of CD34+ cells: 2 x 106 /kg
Success rate: 70-95%
Side effects:
Pain, headaches, arthralgias, malaise, fatigue, insomnia, and nausea
Transient increase in ALP, ALT, LDH, Na and transient decrease in potasium
Increased spleen size in length (in 95% of pts) (mean increase,13%) but 10 days after G-CSF administration, spleen size returns to baseline.
Several cases of spontaneous splenic rupture after G-CSF administration to healthy subjects have been reported.
12
13. --- 2408 unrelated PBSC donors prospectively evaluated by the National Marrow Donor Program (NMDP) between 1999 and 2004.
Incidence of bone pain. Pain symptoms were evaluated before administration of filgrastim each day and at each follow-up after donation. (A) Percentage of PBSC donors who experienced bone pain. (B) Site of bone pain frequency on Day 4. (C) Frequency of highest severity of bone pain during mobilization and collection.
---Pain decreased significantly after filgrastim administration was stopped, but was still reported in 24% and 6% of donors at 2 days and 1 week, respectively, after the final apheresis collection.--- 2408 unrelated PBSC donors prospectively evaluated by the National Marrow Donor Program (NMDP) between 1999 and 2004.
Incidence of bone pain. Pain symptoms were evaluated before administration of filgrastim each day and at each follow-up after donation. (A) Percentage of PBSC donors who experienced bone pain. (B) Site of bone pain frequency on Day 4. (C) Frequency of highest severity of bone pain during mobilization and collection.
---Pain decreased significantly after filgrastim administration was stopped, but was still reported in 24% and 6% of donors at 2 days and 1 week, respectively, after the final apheresis collection.
14. Assessment of donor symptoms using the Abbreviated CALGB Toxicity Criteria. Abbreviated CALGB Toxicity Criteria were evaluated before administration of filgrastim each day. (A) Frequency of highest CALGB score reported by PBSC donors during mobilization and collection. (B) Frequency of highest CALGB score across all symptoms reported by PBSC donors during mobilization and collection.
--Symptoms disappeared almost completely by 1 week after donation, with the exception of slight persisting increases in fatigue, headache, and myalgia (1 week after donation vs baseline: 7.9% vs 0.8%, 4.9% vs 2.5%, and 3.7% vs 2.1%, respectively). Rates of headache and myalgia returned to baseline by the 1-month assessment, but a few donors continued to report fatigue (3.1% at 1 month vs a baseline of 0.8%) Assessment of donor symptoms using the Abbreviated CALGB Toxicity Criteria. Abbreviated CALGB Toxicity Criteria were evaluated before administration of filgrastim each day. (A) Frequency of highest CALGB score reported by PBSC donors during mobilization and collection. (B) Frequency of highest CALGB score across all symptoms reported by PBSC donors during mobilization and collection.
--Symptoms disappeared almost completely by 1 week after donation, with the exception of slight persisting increases in fatigue, headache, and myalgia (1 week after donation vs baseline: 7.9% vs 0.8%, 4.9% vs 2.5%, and 3.7% vs 2.1%, respectively). Rates of headache and myalgia returned to baseline by the 1-month assessment, but a few donors continued to report fatigue (3.1% at 1 month vs a baseline of 0.8%)
15. Box and whiskers plot of blood counts showing the maximum, upper quartile, median, lower quartile, and minimum values obtained from PBSC donors on the first day of injection (Day 1), before and after donation (Day 5 before and Day 5 after), and during follow-up after donation. (A) Donor white blood cell counts. (B) Donor platelet counts. (C) Donor hemoglobin levels, by sex.
----Although nearly a third of donors experienced a WBCs above 50 � 109/L, less than 1% of donors exceeded a higher risk level of 75 � 109/L. No episodes of thrombosis or stroke were reported. After 2 days of collection, nearly 40% of donors had platelet counts below 100 � 109/L, with approximately 2% of donors going below 50 � 109/L and a single donor going below 20 � 109/L. Males and females experienced similar declines in hematocrit values accompanying apheresis, but female donors often declined to values below 30% (5% and 9% occurrence after the first and second apheresis procedure, respectively). Very rarely, donors persisted with low platelet or hemoglobin levels at 1 month after donation-Box and whiskers plot of blood counts showing the maximum, upper quartile, median, lower quartile, and minimum values obtained from PBSC donors on the first day of injection (Day 1), before and after donation (Day 5 before and Day 5 after), and during follow-up after donation. (A) Donor white blood cell counts. (B) Donor platelet counts. (C) Donor hemoglobin levels, by sex.
----Although nearly a third of donors experienced a WBCs above 50 � 109/L, less than 1% of donors exceeded a higher risk level of 75 � 109/L. No episodes of thrombosis or stroke were reported. After 2 days of collection, nearly 40% of donors had platelet counts below 100 � 109/L, with approximately 2% of donors going below 50 � 109/L and a single donor going below 20 � 109/L. Males and females experienced similar declines in hematocrit values accompanying apheresis, but female donors often declined to values below 30% (5% and 9% occurrence after the first and second apheresis procedure, respectively). Very rarely, donors persisted with low platelet or hemoglobin levels at 1 month after donation-
16. G-CSF mobilized vs. BM composition Healthy adult donors:
G-CSF mobilized : 31 donors
BM: 26 donors
16 ---Previous studies by others
have demonstrated that human CD34? cells can be divided into
at least three distinct subsets based on their cell surface
expression of CD45RA and CD123 (IL-3Ra): (i) CD34?
CD45RACD123?/ cells containing common myeloid
progenitors, megakaryocyte/erythrocyte progenitors and more
primitive HSPCs; (ii) CD34?CD45RA?CD123?/ cells containing
more committed granulocyte/macrophage and lymphoid
progenitors; and (iii) CD34dimCD45RA?CD123hi cells.98,99---Previous studies by others
have demonstrated that human CD34? cells can be divided into
at least three distinct subsets based on their cell surface
expression of CD45RA and CD123 (IL-3Ra): (i) CD34?
CD45RACD123?/ cells containing common myeloid
progenitors, megakaryocyte/erythrocyte progenitors and more
primitive HSPCs; (ii) CD34?CD45RA?CD123?/ cells containing
more committed granulocyte/macrophage and lymphoid
progenitors; and (iii) CD34dimCD45RA?CD123hi cells.98,99
17. G-CSF mobilized vs. BM composition 17 BM: minimum of 2 � 108 nucleated cells/kg recipient
PB: minimum 4 x 106
BM: minimum of 2 � 108 nucleated cells/kg recipient
PB: minimum 4 x 106
18. PBSCT compared with BMT PLT engraftment: shorter (~ 5 days)
Neutrophil engraftment: shorter (~ 5 days)
Hospital stay: shorter
Rate of grade II-IV aGVHD: the same
Rate of chronic GVHD: higher (~ 10% higher)
NRM: the same
Relapse rate: the same/trend toward lower rate
OS: the same
BMT CTN Study 0201:
A Phase III Randomized Multicenter Trial Comparing G-CSF Mobilized Peripheral Blood Stem Cell with Marrow Transplantation from HLA Compatible Unrelated Donors
met its target accrual of 550 patients and closed enrollment on September 9, 2009
18
19. Hard to mobilize patients G-CSF non responders: 5-30%
Strategies to manage non-responders:
Dose escalation of G-CSF (12.5�50?�g/kg per day),
Addition of another cytokine such as GM-CSF,
Addition of chemotherapy
Harvesting the BM 19
20. Chemo-mobilization Cyclophosphamide�(typically 2-6 gm/m(2)) plus G-CSF 10 microg/kg on days 3-14
Leukapheresis: After white count recovery (usually days 12-15)
Drawbacks:
higher toxicities,
increased risk for secondary malignances,
impairment of fertility,
cardiac toxicity,
hemorrhagic cystitis,
anaphylactic reactions and higher cost 20 In general, patients who fail initial mobilization are more likely to fail remobilization regardless of the remobilization regimen.17, 25 Furthermore, additional mobilization efforts often result in poorer patient outcomes and increased resource utilization. For these reasons, novel mobilization agents that are less toxic, more rapid and increase the yield of collected CD34+ cells for transplantation are needed.In general, patients who fail initial mobilization are more likely to fail remobilization regardless of the remobilization regimen.17, 25 Furthermore, additional mobilization efforts often result in poorer patient outcomes and increased resource utilization. For these reasons, novel mobilization agents that are less toxic, more rapid and increase the yield of collected CD34+ cells for transplantation are needed.
21. CXCR4/CXCL12 axis
Sinusoidal endothelial cells
1. VCAM-1
2. CXCL12 21 -- CXCL12 or stromal cell-derived factor 1a (SDF-1a) is a chemokine that is constitutively produced at high levels in the BM by stromal cells such as osteoblasts, endothelial cells, Nestin+ MSC, and CAR.
-- It is a potent chemoattractant for HSPCs and has been shown to regulate cell adhesion, survival and cell-cycle status
--- CXCR4 is a member of the large family of seven-transmembrane domain receptors coupled to heterotrimeric Gi proteins and functions as a co-receptor for human immunodeficiency virus type 1 (HIV-1) cell entry
-- The binding of CXCR4 to CXCL12 results in the activation of multiple signal-transduction pathways ultimately triggering chemotaxis -- CXCL12 or stromal cell-derived factor 1a (SDF-1a) is a chemokine that is constitutively produced at high levels in the BM by stromal cells such as osteoblasts, endothelial cells, Nestin+ MSC, and CAR.
-- It is a potent chemoattractant for HSPCs and has been shown to regulate cell adhesion, survival and cell-cycle status
--- CXCR4 is a member of the large family of seven-transmembrane domain receptors coupled to heterotrimeric Gi proteins and functions as a co-receptor for human immunodeficiency virus type 1 (HIV-1) cell entry
-- The binding of CXCR4 to CXCL12 results in the activation of multiple signal-transduction pathways ultimately triggering chemotaxis
22. Genetic alteration of CXCR4/CXCL12 axis CXCR4-/- and CXCL12-/- embryos:
die perinatally
reduced myelopoiesis in fetal liver and nearly complete loss of myelopoiesis in the BM
CXCR4-/- hematopoietic chimeras:
Fetal liver cells from 12- to 15-dpc CXCR4 +/- intercrosses
Transplanted to lethally irradiated mice
22
23. AMD-3100/Plerixafor Reversible inhibitor of CXCR4
Originally tested clinically as an agent for the treatment of HIV infection by blocking the HIV entry into CD4+ T cells
Accidental finding: leukocytosis after a single intravenous dose of plerixafor
A single subcutaneous dose of plerixafor at 160�240?�g/kg: 6- to 10-fold increase in CD34+ cell
Testing of plerixafor as an HIV drug was abandoned:
lack of anti-viral effect
asymptomatic PVC in two patients 23
24. AMD-3100/Plerixafor-Phase I Patients: MM and NHL
Dosage:
160 �g/kg (n = 6)
240 �g/kg (n = 7)
24 --Total WBC count and peripheral blood CD34+ cell count observed in patients receiving AMD3100 at 160 �g/kg or 240 �g/kg at baseline and 4, 6, and 24 hours (WBC count only at 24 hours) after the dose. Each value represents the mean � SE (n = 13).
--Total WBC count and peripheral blood CD34+ cell count observed in patients receiving AMD3100 at 160 �g/kg or 240 �g/kg at baseline and 4, 6, and 24 hours (WBC count only at 24 hours) after the dose. Each value represents the mean � SE (n = 13).
25. AMD-3100/Plerixafor-Phase 2 25 patients with MM or NHL in their first or second CR or PR
G-CSF alone ? 2-week washout period ? AMD3100+G-CSF
G-CSF: 10?�g/kg/day and pheresis was begun on day 4
G-CSF+AMD3100: 160�240?�g/kg/day beginning on day 4 followed 6?h later by
apheresis
Results:
9 of 25 patients who failed to yield the minimum CD34+ cells per kg after
four leukaphereses with G-CSF alone
All 9 patients successfully remobilized with AMD3100+G-CSF
25
26. AMD-3100/Plerixafor-Phase III in NHL Design: phase III, multicenter, randomized (1:1), double-blind, placebo-controlled
Subjects: Patients NHL requiring an Auto-SCT in first or second CR or PR
Study Arms:
G-CSF + AMD3100: 150
G-CSF + Placebo: 148
Primary endpoint:
percentage of patients who collected = 5 � 106 CD34+ cells/kg in < 4 apheresis
Secondary endpoints:
Percantage of patients able to mobilize = 2 � 106 CD34+ cells/kg in = 4 apheresis
the number of apheresis days required to reach = 5 � 106 CD34+ cells/kg;
neutrophil and platelet engraftment
engraftment at 100 days, 6 months, and 12 months after transplantation
26
27. (A) Kaplan-Meier estimate of proportion of patients reaching = 5 � 106 CD34+ cells/kg. The median number of apheresis days required to achieve = 5 � 106 CD34+ cells/kg was 3 days in the plerixafor group, but the median time was not estimable in the placebo group, as less than 50% of patients reached the target within 4 apheresis days. (B) Kaplan-Meier estimate of proportion of patients reaching = 2 � 106 CD34+ cells/kg. Plerixafor-treated patients collected = 2 � 106 CD34+ cells after a median of 1 day of apheresis while placebo-treated patients required a median of 3 days of apheresis. G-CSF, granulocyte colony-stimulating factor; HR, hazard ratio.(A) Kaplan-Meier estimate of proportion of patients reaching = 5 � 106 CD34+ cells/kg. The median number of apheresis days required to achieve = 5 � 106 CD34+ cells/kg was 3 days in the plerixafor group, but the median time was not estimable in the placebo group, as less than 50% of patients reached the target within 4 apheresis days. (B) Kaplan-Meier estimate of proportion of patients reaching = 2 � 106 CD34+ cells/kg. Plerixafor-treated patients collected = 2 � 106 CD34+ cells after a median of 1 day of apheresis while placebo-treated patients required a median of 3 days of apheresis. G-CSF, granulocyte colony-stimulating factor; HR, hazard ratio.
28. Plerixafor-Phase III in MM Design: phase III, multicenter, randomized (1:1), double-blind, placebo-controlled
Subjects: Patients NHL requiring an Auto-SCT in first or second CR or PR
Study Arms:
G-CSF + AMD3100: 148
G-CSF + Placebo: 154
Primary endpoint:
percentage of patients who collected = 6� 106 CD34+ cells/kg in < 2 apheresis
Secondary endpoints:
Percantage of patients able to mobilize = 6 � 106 CD34+ cells/kg in = 4 apheresis
neutrophil and platelet engraftment, and graft failure 28
29. Kinetics of CD34/kg collection. (A) Kaplan-Meier estimate of proportion of patients reaching 6 � 106 or more CD34+ cells/kg. (B) Median CD34+ cells collected on each apheresis day.Kinetics of CD34/kg collection. (A) Kaplan-Meier estimate of proportion of patients reaching 6 � 106 or more CD34+ cells/kg. (B) Median CD34+ cells collected on each apheresis day.
30. Plerixafor in Allo-HSCT 25 HLA-identical sibling donor/recipient pairs
plerixafor 240?�g/kg subcutaneously and underwent leukapheresis 4?h later
first eight patients ? a 10-day washout period ? G-CSF mobilized backup
Median WBC count:
baseline level: 6.76 109/L
4 hours: 18.8 109/L
Median PB CD34+ cells: (median 8-fold increase)
baseline: 2/�L
4 hours:16/�L
Mobilization of >2 � 106 CD34+ cells/kg recipient:
After a single leukapheresis: 2/3
After 2 leukapheresis: 100%
30 --If the minimum CD34 cell dose was not achieved after
the first day of mobilization with AMD3100, the donor was asked to
return in 2 days to repeat the identical process on day 3.
--If the minimum CD34 cell dose was not achieved after
the first day of mobilization with AMD3100, the donor was asked to
return in 2 days to repeat the identical process on day 3.
31. Plerixafor in Allo-HSCT 31 -- The cumulative incidence of grade 2 to 4 acute GVHD was
35%
-- The cumulative incidence of chronic GVHD at 1 year was
33%
-- The cumulative incidence of grade 2 to 4 acute GVHD was
35%
-- The cumulative incidence of chronic GVHD at 1 year was
33%
32. IV Plerixafor Phase I allogeneic transplant trial:
IV plerixafor:80, 160, 240, 320, 400 or 480?�g/kg
After 4 days of drug clearance, SQ 240?�g/kg followed 4?h later by leukapheresis
Peak numbers of circulating CD34+ cells: 4�6?h after IV dosing
320?�g/kg: maximum eightfold increase in circulating CD34+ cells at 4?h after
Phase II study:
28 sibling donors
IV plerixafor : 320?�g/kg followed 4?h later by leukapheresis
leukapheresis yield of 2.0 � 106/kg CD34+ cells/kg:
One leukapheresis: 74%
Two leukopheresis: 95%
32 -- The pharmacokinetics of subcutaneous plerixafor requires that it be administered the night before leukapheresis, so that the morning collection would correspond to the peak of the circulating HSPCs. Such administration is associated with inconvenience and additional cost.
-- Peak numbers of circulating CD34+ cells: 4�6?h after IV dosing (vs 6�9?h after subcutaneous dosing)
-- The pharmacokinetics of subcutaneous plerixafor requires that it be administered the night before leukapheresis, so that the morning collection would correspond to the peak of the circulating HSPCs. Such administration is associated with inconvenience and additional cost.
-- Peak numbers of circulating CD34+ cells: 4�6?h after IV dosing (vs 6�9?h after subcutaneous dosing)
33. Continuous vs. Bolus Plerixafor 33
34. Alternative drugs in development targeting the CXCL12/CXCR4 axis POL6326 (Polyphor, Allschwil, Switzerland)
BKT-140 (4F-benzoyl-TN14003; Biokine Therapeutics, Rehovot, Israel)
NOX-A12 (NOXXON Pharma, Berlin, Germany)
TG-0054 (Taigen Biotechnology, Taipei, Taiwan)
MDX-1338 (BMS-936564; Medarex/Bristol-Myers Squibb, New York, NY, USA)
CXCR4 pepducins (Anchor Therapeutics, Cambridge, MA, USA)
ALX-0651 (Ablynx, Zwijnaarde, Belgium)
AMD070 (Genzyme Corp., Cambridge, MA, USA)
GSK812397 (GlaxoSmithKline, Brentford, England)
KRH-3955 (Kureha Chemical Industries, Nishiki, Japan)
FC131 is a cyclopentapeptide (c(Gly1-D-Tyr2-Arg3-Arg4-Nal5)
WZ811 (N1,N4-di-2-pyridinyl-1,4-benzenedimethanamine) 34
35. VLA-4/VCAM-1 Axis VLA-4 (Very late antigen 4) (a4�1):
an integrin receptor
most leukocytes (monocytes, lymphocytes, eosinophils, basophils) and CD34+ hematopoietic precursor cells
VCAM-1 (vascular cell adhesion molecule 1) or CD106:
Endothelial cells including sinusiodal endothelial cells in vascular niche, CAR cells, Nestin+ MSC, and osteoblasts
Lack of a4 or �1 integrin: embryonic lethal
due to non-hematological defects
Cre/loxP system of conditional a4 ablation:
eightfold increase in the number of circulating
HSPCs (CFU-Cs)
35
36. VLA4/VCAM-1 Axis and VLA-4 Inhibition
Sinusoidal endothelial cells
1. VCAM-1
2. CXCL12 36 Models of HSPC mobilization by G-CSF and inhibitors of CXCR4 and VLA-4. (a) Bone marrow environment at baseline. HSPCs in the
endosteal niche are likely in close contact with osteoblasts and nestin? MSCs, both of which express numerous HSPC retention factors, including
CXCL12, VCAM-1 and SCF. The perivascular niche is more distant from the endosteum and includes both nestin? MSCs and CXCL12 abundant
reticular cells. Cells of the monocytic lineage support the maintenance of osteoblasts and MSCs. b-Adrenergic nerve cells of the sympathetic
nervous system regulate MSC proliferation and induce circadian oscillations of CXCL12 expression. (b) Model of G-CSF-induced HSPC
mobilization. Cells of the monocytic lineage express the receptor for G-CSF and provide factors that support the survival of MSCs and osteoblasts.
Upon 4�5 days of stimulation with G-CSF, the monocytes/macrophages disappear, leading to the loss of osteoblast lineage cells and reduced
expression of CXCL12, VCAM-1 and SCF on MSCs. Reduced expression of these important HSPC retention factors is also observed following
G-CSF signaling through b-adrenergic nerve cells. The net effect of these signaling cascades is the disruption of HSPC retention interactions and
mobilization of HSPCs into the peripheral blood. (c, d) Model of HSPC mobilization by inhibitors of CXCR4 or VLA-4. Targeted disruption of the
interaction of CXCR4 or VLA-4 with their ligands results in the rapid (within hours) and reversible mobilization of HSPCs into the peripheral
circulation. An additive or synergistic affect on HSPC mobilization is observed when a CXCR4 inhibitor is combined with a VLA-4 antagonist,
G-CSF or their combination. When used alone, inhibitors of CXCR4 and VLA-4 mobilize fewer HSPCs than G-CSF.
-----------------------------------------------------
Support for the hypothesis that one cell type is present at various sites within the marrow niche was obtained by a recent landmark study by M�ndez-Ferrer et al. (2010). This group identified a stromal nestin-expressing MSC population (nestin+ MSC) that is closely associated with putative HSCs. Nestin+ MSCs are strictly perivascular and are typically found in more central areas of the marrow, but they are also present in the immediate vicinity of the endosteum, albeit at lower frequency. Moreover, nestin+ MSCs are tightly associated with adrenergic nerve fibers of the sympathetic nervous system (SNS) that regulate HSC mobilization and are responsible for the circadian oscillations in circulating HSC numbers (Katayama et al., 2006; M�ndez-Ferrer et al., 2008). Strikingly, these MSCs express higher levels of HSC maintenance factor transcripts, including CXCL12, stem cell factor (SCF), angiopoietin-1 (Ang-1), IL-7, vascular cell adhesion molecule 1 (VCAM1), and osteopontin (OPN), compared with any other stromal cell type including OBs. Depletion of nestin+ MSCs by inducible expression of diphtheria toxin receptor (DTR) in nestin-expressing cells caused the mobilization of ~50% of HSCs to the spleen. In addition, the homing of transplanted progenitor cells into MSC-depleted recipients was reduced by 90%.Moreover, treatment with the HSC-mobilizing factor granulocyte colony-stimulating factor (G-CSF) decreased the expression of CXCL12 and other factors required for HSC maintenance and retention by nestin+ MSCs. These cells also seem to mediate the capacity of parathyroid hormone to increase HSC numbers, which had previously been correlated with an increase in putative niche OBs (Calvi et al., 2003; Adams et al., 2007). Parathyroid hormone directly stimulated proliferation of nestin+ MSCs while simultaneously promoting their differentiation into OBs (M�ndez-Ferrer et al., 2010). Collectively, these data support the hypothesis that nestin+ MSCs are a functional component of the bone marrow HSC niche.Most interestingly, nestin+ MSCs show several similarities to recently identified mesenchymal progenitors (Sugiyama et al., 2006; Omatsu et al., 2010). These bipotent adipoosteogenic progenitors were identified in a mouse strain in which GFP was expressed from the endogenous CXCL12 locus (Sugiyama et al., 2006; Omatsu et al., 2010). Because of their high CXCL12 expression and their long cellular processes, these cells were named CXCL12-abundant reticular (CAR) cells. The majority of putative HSCs are found in close proximity to CAR cells by immunohistochemistry, and like nestin+ MSCs, CAR cells are predominantly found in the more central areas of the marrow, with some also located close to vessels near the endosteum (Fig. 1). Although CAR cells are more abundant than nestin+ MSCs, they too are tightly associated with sinusoidal endothelium and have a similar morphology to vascular pericytes. These data are in agreement with studies in humans, suggesting that virtually all MSC activity is found within the larger pericyte population that associates closely with the vascular system in the entire body (Crisan et al., 2008).Similar to nestin+ MSCs, CAR cells express HSC maintenance proteins such as CXCL12 and SCF. Induced depletion of CAR cells using a DTR approach causes a partial loss of HSC activity associated with a decrease in HSC cycling, suggesting that CAR cells promote HSC cycling and self-renewal. Even though this phenomenon was not observed after depletion of nestin+ MSCs, it seems likely that CAR cells and nestin+ MSCs represent two highly overlapping CXCL12-expressing cell populations. Because nestin+ MSCs are approximately four times less abundant than CAR cells, contain all colony-forming-unit fibroblast activity within the marrow, harbor high self-renewal activity in vitro and in vivo, and are capable of multilineage differentiation into bone, cartilage, and fat, it appears that nestin+ MSCs may represent a more primitive population compared with CAR cells and may even be a CAR subpopulation. As it remains unclear whether nestin+ MSCs are homogeneous and whether they all express high levels of CXCL12, it is possible that some nestin+ MSCs may not be CAR cells. Although future studies will need to dissect the cellular relationship between these two niche cell populations, there is now convincing evidence that perivascular mesenchymal stem/progenitor cells are critical inhabitants of the HSC niche. (Edinger et al. JEM vol. 208 no. 3 421-428 )
----SNS promotes egress by reducing the expression of key retention factors by the niche cell (M�ndez-Ferrer et al., 2010b), and in contrast, BM Macrophage promotes the expression of these genes and HSC/progenitor retention in the BM.Models of HSPC mobilization by G-CSF and inhibitors of CXCR4 and VLA-4. (a) Bone marrow environment at baseline. HSPCs in the
endosteal niche are likely in close contact with osteoblasts and nestin? MSCs, both of which express numerous HSPC retention factors, including
CXCL12, VCAM-1 and SCF. The perivascular niche is more distant from the endosteum and includes both nestin? MSCs and CXCL12 abundant
reticular cells. Cells of the monocytic lineage support the maintenance of osteoblasts and MSCs. b-Adrenergic nerve cells of the sympathetic
nervous system regulate MSC proliferation and induce circadian oscillations of CXCL12 expression. (b) Model of G-CSF-induced HSPC
mobilization. Cells of the monocytic lineage express the receptor for G-CSF and provide factors that support the survival of MSCs and osteoblasts.
Upon 4�5 days of stimulation with G-CSF, the monocytes/macrophages disappear, leading to the loss of osteoblast lineage cells and reduced
expression of CXCL12, VCAM-1 and SCF on MSCs. Reduced expression of these important HSPC retention factors is also observed following
G-CSF signaling through b-adrenergic nerve cells. The net effect of these signaling cascades is the disruption of HSPC retention interactions and
mobilization of HSPCs into the peripheral blood. (c, d) Model of HSPC mobilization by inhibitors of CXCR4 or VLA-4. Targeted disruption of the
interaction of CXCR4 or VLA-4 with their ligands results in the rapid (within hours) and reversible mobilization of HSPCs into the peripheral
circulation. An additive or synergistic affect on HSPC mobilization is observed when a CXCR4 inhibitor is combined with a VLA-4 antagonist,
G-CSF or their combination. When used alone, inhibitors of CXCR4 and VLA-4 mobilize fewer HSPCs than G-CSF.
-----------------------------------------------------
Support for the hypothesis that one cell type is present at various sites within the marrow niche was obtained by a recent landmark study by M�ndez-Ferrer et al. (2010). This group identified a stromal nestin-expressing MSC population (nestin+ MSC) that is closely associated with putative HSCs. Nestin+ MSCs are strictly perivascular and are typically found in more central areas of the marrow, but they are also present in the immediate vicinity of the endosteum, albeit at lower frequency. Moreover, nestin+ MSCs are tightly associated with adrenergic nerve fibers of the sympathetic nervous system (SNS) that regulate HSC mobilization and are responsible for the circadian oscillations in circulating HSC numbers (Katayama et al., 2006; M�ndez-Ferrer et al., 2008). Strikingly, these MSCs express higher levels of HSC maintenance factor transcripts, including CXCL12, stem cell factor (SCF), angiopoietin-1 (Ang-1), IL-7, vascular cell adhesion molecule 1 (VCAM1), and osteopontin (OPN), compared with any other stromal cell type including OBs. Depletion of nestin+ MSCs by inducible expression of diphtheria toxin receptor (DTR) in nestin-expressing cells caused the mobilization of ~50% of HSCs to the spleen. In addition, the homing of transplanted progenitor cells into MSC-depleted recipients was reduced by 90%.Moreover, treatment with the HSC-mobilizing factor granulocyte colony-stimulating factor (G-CSF) decreased the expression of CXCL12 and other factors required for HSC maintenance and retention by nestin+ MSCs. These cells also seem to mediate the capacity of parathyroid hormone to increase HSC numbers, which had previously been correlated with an increase in putative niche OBs (Calvi et al., 2003; Adams et al., 2007). Parathyroid hormone directly stimulated proliferation of nestin+ MSCs while simultaneously promoting their differentiation into OBs (M�ndez-Ferrer et al., 2010). Collectively, these data support the hypothesis that nestin+ MSCs are a functional component of the bone marrow HSC niche.Most interestingly, nestin+ MSCs show several similarities to recently identified mesenchymal progenitors (Sugiyama et al., 2006; Omatsu et al., 2010). These bipotent adipoosteogenic progenitors were identified in a mouse strain in which GFP was expressed from the endogenous CXCL12 locus (Sugiyama et al., 2006; Omatsu et al., 2010). Because of their high CXCL12 expression and their long cellular processes, these cells were named CXCL12-abundant reticular (CAR) cells. The majority of putative HSCs are found in close proximity to CAR cells by immunohistochemistry, and like nestin+ MSCs, CAR cells are predominantly found in the more central areas of the marrow, with some also located close to vessels near the endosteum (Fig. 1). Although CAR cells are more abundant than nestin+ MSCs, they too are tightly associated with sinusoidal endothelium and have a similar morphology to vascular pericytes. These data are in agreement with studies in humans, suggesting that virtually all MSC activity is found within the larger pericyte population that associates closely with the vascular system in the entire body (Crisan et al., 2008).Similar to nestin+ MSCs, CAR cells express HSC maintenance proteins such as CXCL12 and SCF. Induced depletion of CAR cells using a DTR approach causes a partial loss of HSC activity associated with a decrease in HSC cycling, suggesting that CAR cells promote HSC cycling and self-renewal. Even though this phenomenon was not observed after depletion of nestin+ MSCs, it seems likely that CAR cells and nestin+ MSCs represent two highly overlapping CXCL12-expressing cell populations. Because nestin+ MSCs are approximately four times less abundant than CAR cells, contain all colony-forming-unit fibroblast activity within the marrow, harbor high self-renewal activity in vitro and in vivo, and are capable of multilineage differentiation into bone, cartilage, and fat, it appears that nestin+ MSCs may represent a more primitive population compared with CAR cells and may even be a CAR subpopulation. As it remains unclear whether nestin+ MSCs are homogeneous and whether they all express high levels of CXCL12, it is possible that some nestin+ MSCs may not be CAR cells. Although future studies will need to dissect the cellular relationship between these two niche cell populations, there is now convincing evidence that perivascular mesenchymal stem/progenitor cells are critical inhabitants of the HSC niche. (Edinger et al. JEM vol. 208 no. 3 421-428 )
----SNS promotes egress by reducing the expression of key retention factors by the niche cell (M�ndez-Ferrer et al., 2010b), and in contrast, BM Macrophage promotes the expression of these genes and HSC/progenitor retention in the BM.
37. BIO5192, a small molecule inhibitor of VLA-4 37 BIO 5192: Peripheral blood CFU-GM levels peaked 0.5�1?h after BIO5192 administration and returned to baseline by 6?h.
=== Here we show that interruption of the
VCAM-1/VLA-4 axis with a small molecule
inhibitor of VLA-4, BIO5192, results in a
30-fold increase in mobilization of murine
hematopoietic stem and progenitors
(HSPCs) over basal levels. An additive
affect on HSPC mobilization (3-fold) was
observedwhenplerixafor (AMD3100), a small
molecule inhibitor of the CXCR-4/SDF-1
axis, was combined with BIO5192. Furthermore,
the combination of granulocyte
colony-stimulating factor (G-CSF),
BIO5192, and plerixafor enhanced mobilization
by 17-fold compared with G-CSF
alone. HSPCs mobilized by BIO5192 or
the combination of BIO5192 and plerixafor
mobilized long-term repopulating
cells, which successfully engraft and expand
in a multilineage fashion in secondary
transplantation recipients. Splenectomy
resulted in a dramatic enhancement
of G-CSF�induced mobilization while decreasing
both plerixafor- and BIO5192-
induced mobilization of HSPCs. These
data provide evidence for the utility of
small molecule inhibitors of VLA-4 either
alone or in combination with G-CSF or
AMD3100 for mobilization of hematopoietic
stem and progenitor cells. (Blood.
2009;114:1340-1343)
===========
Natalizumab (Tysabri; Biogen/Idec, Cambridge, MA, USA) is a recombinant humanized neutralizing IgG4 monoclonal antibody that binds to the a4-subunit of the a4�1 (VLA-4) and a4�7 integrins. Overall, the number of circulating CD34+ cells per �l blood increased three- to fivefold during the first 72?h after treatment and remained elevated at these levels (8�10 CD34+ cells per �l) for at least a month after natalizumab injection. Somewhat surprisingly, repeated administration of the antibody failed to mobilize additional CD34+ cells.
===
As BIO5192 will not be clinically developed, we have performed identical studies with similar results using the small-molecule VLA-4 antagonist firategrast, which is currently in clinical development for the treatment of MS by GlaxoSmithKline (data not shown).BIO 5192: Peripheral blood CFU-GM levels peaked 0.5�1?h after BIO5192 administration and returned to baseline by 6?h.
=== Here we show that interruption of the
VCAM-1/VLA-4 axis with a small molecule
inhibitor of VLA-4, BIO5192, results in a
30-fold increase in mobilization of murine
hematopoietic stem and progenitors
(HSPCs) over basal levels. An additive
affect on HSPC mobilization (3-fold) was
observedwhenplerixafor (AMD3100), a small
molecule inhibitor of the CXCR-4/SDF-1
axis, was combined with BIO5192. Furthermore,
the combination of granulocyte
colony-stimulating factor (G-CSF),
BIO5192, and plerixafor enhanced mobilization
by 17-fold compared with G-CSF
alone. HSPCs mobilized by BIO5192 or
the combination of BIO5192 and plerixafor
mobilized long-term repopulating
cells, which successfully engraft and expand
in a multilineage fashion in secondary
transplantation recipients. Splenectomy
resulted in a dramatic enhancement
of G-CSF�induced mobilization while decreasing
both plerixafor- and BIO5192-
induced mobilization of HSPCs. These
data provide evidence for the utility of
small molecule inhibitors of VLA-4 either
alone or in combination with G-CSF or
AMD3100 for mobilization of hematopoietic
stem and progenitor cells. (Blood.
2009;114:1340-1343)
===========
Natalizumab (Tysabri; Biogen/Idec, Cambridge, MA, USA) is a recombinant humanized neutralizing IgG4 monoclonal antibody that binds to the a4-subunit of the a4�1 (VLA-4) and a4�7 integrins. Overall, the number of circulating CD34+ cells per �l blood increased three- to fivefold during the first 72?h after treatment and remained elevated at these levels (8�10 CD34+ cells per �l) for at least a month after natalizumab injection. Somewhat surprisingly, repeated administration of the antibody failed to mobilize additional CD34+ cells.
===
As BIO5192 will not be clinically developed, we have performed identical studies with similar results using the small-molecule VLA-4 antagonist firategrast, which is currently in clinical development for the treatment of MS by GlaxoSmithKline (data not shown).
38. Summary PBSC is the main source of stem cell for HSCT
G-CSF has 5-30% failure rate in mobilization
Strategies to manage hard to mobilize patients:
Dose escalation of G-CSF (12.5�50?�g/kg per day),
Addition of another cytokine such as GM-CSF,
Addition of chemotherapy
Harvesting the BM
CXCR4 and VLA-4 are the dominant receptors regulating HSPC migration and retention within the BM
Plerixafor in combination with G-CSF, is FDA approved for HSPC mobilization in patients with NHL and MM
VLA-4 inhibitors either alone or in combination with G-CSF or plerixafor increase mobilization yield.
Dual inhibitor approach may ultimately provide a more efficient method to collect HSC in a single day
38
39. References 1. www.NMPD.org
2. Iskra Pusic and John F. DiPersio. Current Opinion in Hematology 2010,17:319�326
3. Wilson et al. Bone-marrow haematopoietic-stem-cell niches. Nat Rev Immunol 2006; 6: 93�106
4. MP Rettig, G Ansstas and JF DiPersio. Leukemia. 2011 Sep 2. doi: 10.1038/leu.2011.197. [Epub ahead of print]
5. Christopher M et al. J Exp Med 2011 208:251-260
6. McCullough et al.. Transfusion 2008;48(9):2008-2025
7. Pulsipher M A et al. Blood 2009;113:3604-3611
8. Hassan et al. Transplant Immunology, 1996; 4: 319-323
9. Blais et al. J Clin Oncol 2000;18(3):537-546
10. Powles et al. Lancet 2000;355(9211):1231-1237
11. Ringden et al. J Clin Oncol 2002;20(24):4655-4664
12. Bensinger et al. N Engl J Med 2001;344(3):175-181
13. Kos et alJ Clin Oncol. 2000 May;18(9):1824-30
14. De Clercq E. The bicyclam AMD3100 story. Nat Rev Drug Discov 2003; 2: 581�587
15. Devine S M et al. JCO 2004;22:1095-1102
16. Flomenberg et al. Blood 2005; 106: 1867�1874
17. DiPersio J F et al. JCO 2009;27:4767-4773
18. DiPersio J F et al. Blood 2009;113:5720-5726
19. Devine S M et al. . Blood 2008; 112: 990�998
20. Bonig H et al. Experimental Hematology 2009;37:402�415
21. Scott LM et al. Mol Cell Biol 2003; 23: 9349�9360
22. Ramirez P et al. Blood. 2009;114:1340-1343
39