Download
1 / 16
160 likes | 385 Vues
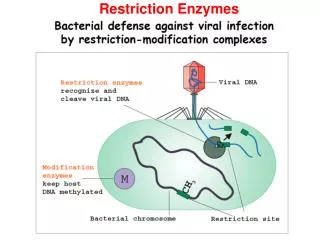
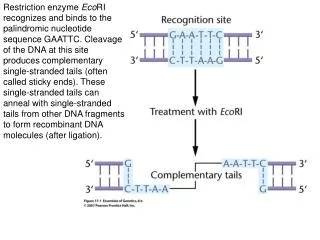
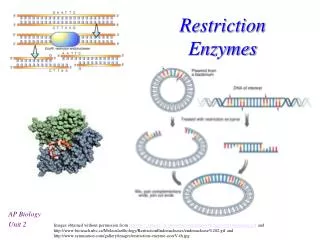
Restriction Enzymes. Lecture 16: 1 11/29/2006. 10. Activities of some restriction enzymes are methylation-dependent. CH3 5’……GATC……...3’ 5’……CTAG……...3’ CH3 This property is exploited to develop a site-directed mutagenesis

E N D
Restriction Enzymes Lecture 16: 1 11/29/2006 10. Activities of some restriction enzymes are methylation-dependent. CH3 5’……GATC……...3’ 5’……CTAG……...3’ CH3 This property is exploited to develop a site-directed mutagenesis (Quickchange Site-Directed Mutagenesis kit from Stratagene). (See transparency) 11. RFLP and RAPD Restriction Fragment Length Polymorphism (RFLP) Restriction Fragment Length Polymorphism (RFLP) is a technique in which organisms may be differentiated by analysis of patterns derived from cleavage of their DNA. If two organisms differ in the distance between sites of cleavage of a particular restriction endonuclease, the length of the fragments produced will differ when the DNA is digested with a restriction enzyme. The similarity of the patterns generated can be used to differentiate species (and even strains) from one another. cystic fibrosis (CF)
Restriction Enzymes Lecture 16: 2 11/29/2006 • Random amplified polymorphic DNAs (RAPDs): Random primers are used to amplify DNA fragments from a DNA template by PCR. The resulting PCR products are subsequently digested with restriction enzymes and analyzed on an agarose gel. The pattern of restriction fragments can then used to reveal the genetic differences between individuals or populations. • 12. DNA methlyase: enyzymes that transfer a methyl group from S-adenossylmethionine to either adenine or cytosine residues. • Most of laboratory strains of E. coli contain three site-specific DNA methylases. • Dam methylase (DNA adenine methylase) transfers a methyl group to the N6 position of the adenine residues in the sequence GATC. • 2) DCM methylates (DNA cytosine methylase) the internal cytosine residues in the sequences CCAGG and CCTGG at the C5 position. • The EcoK I methylase modifies adenine residues in the sequences AAC(N6)GTGC and GCAC(N6)GTT. • Many laboratory E. coli strains lack Dam and Dcm. • In higher eukaryotes, CpG methlyases transfer a methyl group to the C5 poisition of cytosine residues.Patterns of CpG methylation are heritable, tissue specific, and correlate with gene expression. • 13. If the restriction sites are located at the ends of the DNA fragment, several extra nucleotides need to be added to it for efficient digestions by restriction enzymes. • 14. Spermidine [the cationic polyamines spermidine {[H3N(CH2)3NH2(CH2)4NH3]3+}] (~5 mM) can be included to help digest recalcitrant genomic DNA.
DNA Replication Lecture 16: 3 11/29/2006 DNA Replication is Semiconservative
DNA Replication Lecture 16: 4 11/29/2006 Dr. Mattew Meselson and his recent research on asexual organsims The bdelloid rotifer Philodina roseola (ca 400 microns); her ancestors abandoned sexual reproduction 80-100 million years ago.
DNA Replication Lecture 16: 5 11/29/2006 They labelled bacterial chromosomes uniformly with the 15N or "heavy" isotope of nitrogen by growing Escherichia coli cells in the presence of 15NH4Cl for a number of generations. They harvested the cells, washed them free of the labelled medium and then allowed the cells to grow in a medium containing 14NH4Cl -- i.e. the "light" isotope of nitrogen. They then analysed the DNA and the dilution of the heavy isotope as the cells replicated. Their analysis made us of CsCl equilibrium density gradients -- one of the first uses of this technique. Meselson and Stahl needed to follow the cells through at least two rounds of replication. They disproved the conservative model of replication after observing and analyzing a single round of replication, but a second round of replication was necessary to disprove the dispersive model of DNA replication.
DNA Replication Lecture 16: 6 11/29/2006 Evidence for bidirectional replication
DNA Replication Lecture 16: 7 11/29/2006 Evidence for bidirectional replication John Cairns provided a very elegant demonstration of E. coli chromosomal replication in 1963. He radioactively labelled the chromosome by growing his cultures in a medium containing 3H-thymidine. The nucleoside base was incorporated uniformly into the bacterial chromosome. He then isolated the chromosomes by lysing the cells very very gently and placed them on an electron micrograph (EM) grid which he exposed to X-ray film for two months. The replication intermediates first described by Cairns are known as theta forms due to their resemblance to the greek letter theta - q. They are also known as Cairns forms. Cairns showed that: The E. coli chromosome is circular. This had been suspected from genetic mapping evidence but direct physical confirmation was a nice demonstration that the genetic results were valid. There is a single origin of replication in E. coli.. However, Cairns result were also open to some interpretation: His results did not allow one to discriminate between unidirectional or bidirectional modes of replication. His results were consistent with either model. The location of the origin of replication could not be determined as such. Although, it was clear that each chromosome had a single origin of replication, this did not establish whether every chromosome had exactly the same origin of replication.
DNA Replication Lecture 16: 8 11/29/2006 Evidence for bidirectional replication E.coli cells are exposed first to high and then to low concentrations of [3H]thymine, the resulting DNA will be heavily labeled near replication origins.
DNA Replication Lecture 16: 9 11/29/2006 Evidence for bidirectional replication
DNA Replication Lecture 16: 10 11/29/2006 Replication origins E. coli Replication Origin OriC. OriC is an 240-bp DNA fragment that contains repetitive 9-bp and AT-rich 13-bpsequences, referred to as 9-mers (dna Boxes) and 13 mers, respectively. Yeast autonomously sequences (ARS): ARS is a sequence of 180 bp that confers on a plasmid the ability to replicate in yeast. ARS contains a 15-bp segment essential for replication with a consensus, 5’ a/t-TTTAT-a/g-TTT-a/t 3’. SV40 Replication origin: a 65-bp DNA fragment.
DNA Replication Lecture 16: 11 11/29/2006 In the 1950s, Arthur Kornberg recognized that the energetics of the biochemical synthesis of a polynucleotide chain would require deoxynucleoside triphosphates (dNTPs). He also realized that he needed a very sensitive assay for DNA synthesis and that if he radioactively labelled the a phosphate of dNTPs he could monitor the incorporation of the labelled dNTPs into DNA with a simple assay (actually precipitation with trichloroacetic acid - TCA). By choosing E. coli for his studies, he was also able to prepare large quantities of cells and to obtain large amounts of purified enzyme. In this way he discovered an enzyme in E. coli that could catalyse the synthesis of DNA. He called it DNA polymerase. We now know it as DNA polymerase I. We also now know that there are four other DNA polymerase enzymes in E. coli.
DNA Replication Lecture 16: 12 11/29/2006 Why can DNA polymerase only act from 5' to 3'? The reason is the relative stability of each end of DNA. A triphosphate is required to provide energy for the bond between a newly attached nucleotide and the growing DNA strand. However, this triphosphate is very unstable and can easily break into a monophosphate and an inorganic pyrophosphate, which floats away into cell. At the 5' end of the DNA, this triphosphate can easily break, so if a strand has been sitting in the cell for a while, it would not be able to attach new nucleotides to the 5' end once the phosphate had broken off. On the other hand, the 3' end only has a hydroxyl group, so as long as new nucleotide triphosphate are always brought by DNA polymerase, synthesis of a new strand can continue no matter how long the 3' end has remained free. This presents a problem, since one strand of the double helix is 5' to 3' , and the other one is 3' to 5'. How can DNA polymerase synthesize new copies of the 5' to 3' strand, if it can only travel in one direction? This strand is called the lagging strand, and DNA polymerase makes a second copy of this strand in spurts, called Okazaki fragments, as shown in the diagram. The other strand can proceed with synthesis directly, from 5' to 3', as the helix unwinds. This is the leading strand.
DNA Replication Lecture 16: 13 11/29/2006 E. coli DNA polymerase I DNA polymerase I is a 928 amino-acid polypeptide (MW=103118) encoded by the polA gene. It has three distinct enzymatic activities: A 5' -> 3' polymerase activity A 3' -> 5' exonuclease activity A 5' -> 3' exonuclease activity Structurally, the enzyme has two domains which can be separated by proteolysis with trypsin or subtilisin, both of which cleave at the amino acid 324. The smaller (34 kD) N-terminal fragment carries the 5' -> 3' exonuclease activity. The larger (76 kD) C-terminal fragment carries the polymerase and 3' -> 5' exonuclease activities. This fragment is often called Klenow fragment after Hans Klenow who first discovered this proteolytic cleavage. The function of the 3' -> 5' exonuclease activity is that of PROOF-READING. Any nucleotides -- such as transiently pairing tautomers -- that are incorrectly incorporated are excised by this activity.
DNA Replication Lecture 16: 14 11/29/2006 • DNA polymerase I is not the replicative polymerase. • Until 1969, DNA polymerase I was the only known DNA polymerase in E. coli. However, as DNA polymerase I was characterized further, it became clear that the properties of this enzyme were not suitable for an enzyme with a central role in DNA replication: • The enzyme is too slow!DNA polymerase I catalyzes the incorporation of dNTPs at a maximal rate of 20 nt/sec (approximately). At this rate, it would require approximately 460,000 sec (= 7667 min = 128 hr = 5.3 days) to replicate the entire E. coli chromosome! This is much too slow for an organism which can divide every 20 mins. • The enzyme is too abundantThere are approximately 400 molecules of DNA polymerase I per E. coli cell. This is excessive given that there are generally only 2 replication forks per cell. • The enzyme is not processive enoughDNA polymerase I dissociates after catalysing the incorporation of 20-50 nucleotides. • DNA polymerase I cannot initiate DNA synthesis de novo.However, it must be noted that DNA polymerase I is not unique in this regard. It shares this particular problem with every other known DNA polymerase. • Cells containing the polA1 mutation are viable.In 1969, De Luca and Cairns reported the isolation of a mutant of E. coli that was viable even though it lacked DNA polymerase I activity. The mutant was found to be an amber mutant; we now know from sequence information that the codon for tryptophan 342 is mutated to a stop codon in this mutant.Cells carrying this mutation grow at normal rates. However, they are more sensitive than normal to mutagenic agents such as UV light. • Because the polA1 mutants grow normally, they were used as in the search for other DNA polymerase activities in E. coli. Two were found: DNA polymerase II and DNA polymerase III.
DNA Replication Lecture 16: 15 11/29/2006 DNA polymerase II This enzyme is most likely involved in DNA repair systems. The enzyme is 89921 kD in size and is coded by the polB gene. Strains lacking the gene show no defect in growth or replication. Synthesis of PolII is induced during the stationary phase of cell growth. This is a phase in which little growth and DNA synthesis occurs. It is also a phase in which the DNA can accumulate damage such as short gaps, which act as a block to PolIII. Under these circumstances, PolII helps to overcome the problem because it can reinitiate DNA synthesis downstream of gaps. PolII has a low error rate but it is much too slow to be of any use in normal DNA synthesis.