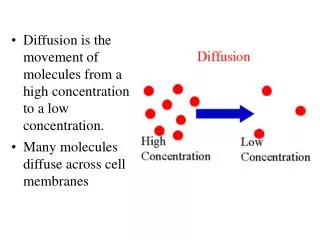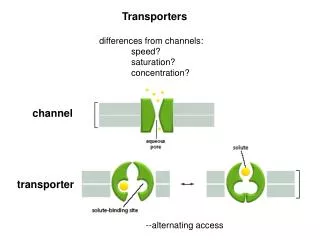
Transporters differences from channels: speed? saturation? concentration?
360 likes | 641 Vues
Transporters differences from channels: speed? saturation? concentration?. channel. transporter. --alternating access. transporters pumps ionic coupling. Pumps. pumps generate the ionic gradients that drive flux through channels and through other transporters

Transporters differences from channels: speed? saturation? concentration?
E N D
Presentation Transcript
Transporters differences from channels: speed? saturation? concentration? channel transporter --alternating access
transporters pumps ionic coupling Pumps pumps generate the ionic gradients that drive flux through channels and through other transporters energy provided by ATP hydrolysis relatively small number drive flux at plasma membrane secretory pathway (including endocytic) mitochondria
P-type ATPase: Na+/K+-ATPase phosphorylated intermediate Na/K-ATPase makes Na+, K+ gradients H/K-ATPase acidifies stomach phospholipid flippases (maintain asymmetry of bilayer) Ca/H-ATPase removes cytosolic Ca++
how do ATP binding, phosphorylation drive flux? crystallization of Ca++-ATPase --in multiple conformations acidic residues bind ions two gates never open together both gates close when ion bound N, P, A control gates each event triggered by previous (Olesen et al, 2007)
Rotary Club: F0F1 ATPase (ATP synthase) matrix (Junge et al., 2009) cyto H+ flux drives unidirectional rotation and ATP synthesis
V-type H+-ATPase V-type F0F1 uses ATP to make H+ electrochemical gradient in endosomes, lysosomes, synaptic vesicles 3 ATP/rotation (at A/B interface) (both V- and F0F1) what confers difference in direction?
transporters pumps ionic coupling secondary active transport
big conformational changes as well as faster binding, unbinding movement of unloaded carrier essential for net flux movement of unloaded carrier must be distinct from loaded
exchange reaction --no net flux
membrane vesicles were loaded with 14C-glucose diluted into medium containing different concentrations of non-radioactive glucose (indicated on the x-axis) *glucose glucose avoids movement of unloaded carrier translocation steps slow --channel cannot do this heteroexchange obligate exchangers cannot do net flux amphetamines may release monoamines this way (exchange-diffusion)
dopamine transporter striatal slice voltammetry stimulation impaired dopamine clearance 95% decrease in dopamine stores! --role in recycling as well as clearance
coupling rules required for active transport--why? AND operation
So + Na+o Si + Na+i • stoichiometry of 1 Na+ : 1 S and ~12-fold Na+ gradient • will generate ? gradient of S • at equilibrium, equal rates in and out of cell • [Na+]o x [S]o = [Na+]i x [S]i • [Na+]o / [Na+]i = [S]i / [S]o • what if it is an exchanger?
2 Na+ 2 Na+ So + 2Na+o Si + 2Na+i • if coupling involves 2 Na+ : 1 S, then • [Na+]o2 x [S]o = [Na+]i2 x [S]i • ([Na+]o / [Na+]I)2 = [S]I / [S]o or • log10 (Sin/Sout) = 2 log10 (Na+out/Na+in) • why not just make the stoichiometry very high? • and what if net flux involves charge movement?
electrogenic transport (transport that moves net charge) • the negative resting membrane potential • augments the chemical gradient for Na+ by • – zTVm/ 60 mV (log units) • where zT = net charge moved n Na+ n Na+ • added to the concentration gradient, • log10 (Sin/Sout) = n log10 (Na+out/Na+in) – zT/ 60 mV • where n = # Na+ ions cotransported • -- the power of membrane potential! • this equation changes for different ionic coupling
glycine transport nNa+ gly Cl- electrogenic transport produces currents --depend on Na+, Cl- defined by gly addition can measure charge:flux using labeled glycine, Cl (Roux and Supplisson, 2000)
why determine coupling? ionic coupling determines direction of flux magnitude of gradient (can exceed 106:1) regulation by membrane potential • for electrogenic glycine transport, • log10 (glyin/glyout) = m log10 (Na+out/Na+in) + n log10 (Cl-out/Cl-in) – zT/ 60 mV • zT/ 60 mV = log10 Na+om x Cl-n x So • Na+im x Cl-in x Si • = 60 mV log10 Na+om x Cl-i n x So = Erev • zT Na+im x Cl-in x Si • --like Nernst equation: • ENa = 60 mV log10 Na+o/Na+i • what are the differences?
GlyT1 GlyT2 reversal potentials used to determine stoichiometry
(Gomeza et al, 2003) neuron glia GlyT1 KO: excess glycine (excess inhibition)--main role clearance GlyT2 KO: resembles GlyR KO (startle)--main role packaging differences in ionic coupling can also confer transfer between cells
excitatory amino acid transporters (EAATs) bacterial transporter trimer (bowl): 3 Na+:1 H+:1 glu- 1 K+ (Boudker et al, 2007) stoichiometry predicts huge gradients control activation of perisynaptic receptors, spillover
transport within monomer HP1,2 the gates? structure in out and inward conformations rigid body motion (Reyes et al., 2009)
3 Na+:1 H+:1 glu- 1 K+ glutamate added to the outside must trigger inward + charge movement glu uncoupled uncoupled (Wadiche et al, 1995) glutamate-induced currents reverse for EAAT1 and 3 --glutamate-gated chloride channel (receptor)
some transporters also behave like channels transport cycle can gate an ion channel ?evolutionary intermediate
Shaker voltage-gated K+ channel R365H no Na+ or K+ (~gating charge) depends on H+ current flow in direction of pH gradient --voltage sensor turned into transporter (Starace et al., 1997)
+ + + + + + + + + + + Cl- H+concentrations much lower than other ions gradients easier to make, break --can also be regulated, such as membrane potential without DpH!! (mito have Vm ~ -160 mV, DpH < 0.5) ATP H+ ADP H+ H+ H+ H+ --acridine orange quenches when trapped in acidic membranes: H+ SV in absence of Cl-, H+-ATPase makes only membrane potential --Cl- entry required for acidification --DpH, DY can be regulated independently
+ + + Cl- ClC family includes Cl- channels and Cl-/H+ exchangers intracellular ClCs mediate exchange of 2Cl- : 1 H+ why couple to H+ exchange, which ought to dissipate DpH? ATP ATP H+ H+ ADP ADP + H+ H+ H+ 2 Cl- H+ exchange mechanism makes bigger DpH?!
several crystal structures ClC transporter is a channel with a lever arm --can count Cl- ions?! --Cl- itself a gate --alternating access without large conformational change (Feng et al., 2010)
what confers the progressive acidification of endosomes? (Faundez and Hartzell, 2004) same stoichiometry of coupling (2 Cl- : 1 H+) predicts same DpH same cytosolic [Cl]
+ + + + H+ H+ + H+ H+ + + + + + is there an opposing activity that dissipates DpH? ATP H+ ADP H+ Na+/K+ plasma membrane Na+/H+ exchangers regulate cell pH intracellular isoforms mediate K+/H+ exchange, dissipate DpH mechanism to make Dy
Differences from Channels 1) Alternating access rather than pore importance of unloaded carrier for net flux (active transport) exchange 2) Pumps use ATP to make ionic gradients 3) Ionic coupling uses these gradients to drive movement of other solutes behavior not predicted by Nernst equation direction of flux, stoichiometry can vary reversal potential can be used to determine coupling coupling dictates physiological role 4) Ions interact more tightly than with channel 5) Transport cycle can gate channel (evolution of channel gating?) 6) Bulk H+ concentrations can also change (like Ca++) DpH promoted by anion entry DY promoted by cation entry
Background Alberts, Chapter 11 Nicholls, D.G. and Ferguson, S.J. Bioenergetics 2, Academic Press, 1992. Stein, W. Channels, Carriers and Pumps. Academic Press, 1990. References Accardi, A., and Miller, C. (2004). Secondary active transport mediated by a prokaryotic homologue of ClCCl- channels. Nature 427, 803-807. Dallos, P., and Fakler, B. (2002). Prestin, a new type of motor protein. Nat Rev Mol Cell Biol 3, 104-111. Goh, G.,Y. Huang, H., Ullman, J., Borre, L., Hnasko, T.S., Trussell, L.O. and Edwards, R.H. 2011. Presynaptic regulation of quantal size: K+/H+ exchange stimulates glutamate storage by increasing membrane potential. Nat. Neurosci. 14, 1285-1292. Feng, L., Campbell, E.B., Hsiung, Y., and MacKinnon, R. (2010). Structure of a eukaryotic CLC transporter defines an intermediate state in the transport cycle. Science 330, 635-641. Gomeza, J., Hulsmann, S., Ohno, K., Eulenburg, V., Szoke, K., Richter, D., and Betz, H. (2003). Inactivation of the glycine transporter 1 gene discloses vital role of glial glycine uptake in glycinergic inhibition. Neuron 40, 785-796. Gomeza, J., Ohno, K., Hulsmann, S., Armsen, W., Eulenburg, V., Richter, D.W., Laube, B., and Betz, H. (2003). Deletion of the mouse glycine transporter 2 results in a hyperekplexia phenotype and postnatal lethality. Neuron 40, 797-806. Guan, L., and Kaback, H.R. (2006). Lessons from lactose permease. Annu Rev BiophysBiomolStruct 35, 67-91. Hnasko, T.S., Chuhma, N., Zhang, H., Goh, G.A., Sulzer, D., Palmiter, R.D., Rayport, S. and Edwards, R.H. 2010. Vesicular glutamate transport promotes dopamine storage and glutamate coreleasein vivo. Neuron 65, 643-656. Junge, W., Sielaff, H., and Engelbrecht, S. (2009). Torque generation and elastic power transmission in the rotary F(O)F(1)-ATPase. Nature 459, 364-370. Olesen, C., Picard, M., Winther, A.M., Gyrup, C., Morth, J.P., Oxvig, C., Moller, J.V., and Nissen, P. (2007). The structural basis of calcium transport by the calcium pump. Nature 450, 1036-1042. Reyes, N., Ginter, C., Boudker, O. (2009) Transport mechanism of a bacterial homologue of glutamate transporters. Nature 462:880-885. Roux, M.J., and Supplisson, S. (2000). Neuronal and glial glycine transporters have different stoichiometries. Neuron 25, 373-383.
Starace, DM, Stefani, E, Bezanilla, F (1997) Voltage-dependent proton transport by the voltage sensor of the Shaker K+ channel. Neuron 19:1319-1327. Wadiche, J.I., Amara, S.G., and Kavanaugh, M.P. (1995). Ion fluxes associated with excitatory amino acid transport. Neuron 15, 721-728. Wu, Y., Wang, W., Diez-Sampedo, A., Richerson, G.B. (2007) Nonvesicular inhibitory neurotransmission via reversal of the GABA transporter GAT-1. Neuron 56:851-865. Yamashita, A., Singh, S.K., Kawate, T., Jin, Y., and Gouaux, E. (2005). Crystal structure of a bacterial homologue of Na(+)/Cl(-)-dependent neurotransmitter transporters. Nature 437, 215-223. Zerangue, N., and Kavanaugh, M.P. (1995). Flux coupling in a neuronal glutamate transporter. Nature 383, 634-637.