Download

1 / 33
340 likes | 417 Vues
Explore the effector functions of antibodies in neutralizing microbes and toxins, the role of Fcγ receptors in immune regulation, and the impact of complement deficiencies on infection susceptibility. Learn about neonatal immunity and the transfer of maternal antibodies for early protection.

E N D
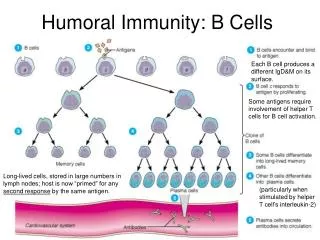
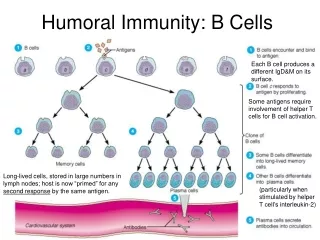
Chapter 13 Effector Mechanisms of Humoral Immunity
Inhibitory Signaling by the FcγRIIB Receptor • Immune complex–mediated cross-linking of the inhibitory FcγRIIB leads to tyrosine phosphorylation of the ITIM in the cytoplasmic tail • One somewhat empirical but often useful treatment of many autoimmune diseases is the intravenous administration of pooled human IgG (IVIG). IVIG may engage FcγRIIB to deliver inhibitory signals to B lymphocytes and other cells, thus reducing antibody production and dampening inflammation
Role of Fcγ Receptors in Phagocytosis andActivation of Phagocytes
Antibody-Mediated Clearance of Helminths • Antibodies, mast cells, and eosinophils function with antibodies to mediate the expulsion and killing of some helminthic parasites • IgE, IgG, and IgA antibodies that coat helminths can bind to Fc receptors on eosinophils and cause the degranulation of these cells, releasing the basic protein and other eosinophil granule contents that kill the parasites • The high affinity Fcε receptor of eosinophils (FcεRI) lacks the signaling β chain and can only signal relatively weakly through the associated γ chain
The early steps of complement activation by the alternative, classical, and lectin pathways
Late steps of complement activation and formation of the MAC
Complement Deficiencies • Genetic deficiencies in classical pathway components, including C1q, C1r, C4, C2, and C3; C2 deficiency is the most common human complement deficiency • More than 50% of patients with C2 and C4deficiencies develop systemic lupus erythematosus • Deficiency of C3 is associated with frequent serious pyogenic bacterial infections that may be fatal, illustrating the central role of C3 in opsonization, enhanced phagocytosis, and destruction of these organisms • Deficiencies in components of the alternative pathway, including properdin and factor D, result in increased susceptibility to infection with pyogenic bacteria
Deficiencies in the terminal complement components, including C5, C6, C7, C8, and C9 disseminated infections by Neisseria bacteria, including Neisseria meningitidis and Neisseria gonorrhoeae • Deficiencies in complement regulatory proteins are associated with abnormal complement activation and a variety of related clinical abnormalities • Deficiencies in complement receptors include the absence of CR3 and CR4, both resulting from rare mutations in the β chain (CD18) gene common to the CD11CD18 family of integrin molecules is characterized by recurrent pyogenic infections and is caused by inadequate adherence of neutrophils to endothelium
Pathologic Effects of a Normal Complement System • Evasion of Complement by Microbes • Microbes can evade the complement system by recruiting host complement regulatory proteins • Some pathogens, like schistosomes, Neisseria gonorrhoeae, and certain Haemophilus species, scavenge sialic acids from the host and enzymatically transfer the sugar to their cell surfaces • GP41 on human immunodeficiency virus (HIV) can bind to factor H, and this property of the virus is believed to contribute to virion protection
NEONATAL IMMUNITY • Neonatal mammals are protected from infection by maternally produced antibodies transported across the placenta into the fetal circulation and by antibodies in ingested milk transported across the gut epithelium of newborns by a specialized process known as transcytosis • Maternal IgG is transported across the placenta, and maternal IgA and IgG in breast milk are ingested by the nursing infant. The transepithelial transport of maternal IgA into breast milk depends on the poly Ig receptor
Transport of maternal IgG across the placenta and across the neonatal intestinal epithelium is mediated by an IgG-specific Fc receptor called the neonatal Fc receptor (FcRn) • The FcRn is unique among Fc receptors in that it resembles a class I MHC molecule containing a transmembrane heavy chain that is noncovalently associated with β2- microglobulin