Photosynthesis
Photosynthesis. This version of the file contains very few pictures – less pretty but takes up MUCH less disc space and download time. To see the images in their full glory, make sure you come to the lecture!!. Introduction.


Photosynthesis
E N D
Presentation Transcript
Photosynthesis This version of the file contains very few pictures – less pretty but takes up MUCH less disc space and download time. To see the images in their full glory, make sure you come to the lecture!!
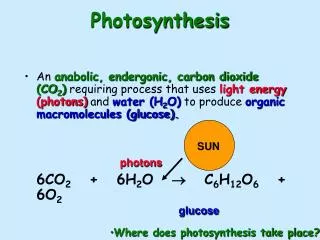
Introduction Photosynthesis is undoubtedly one of the most important processes on the planet. Arguably second to DNA replication (since DNA came first and drives photosynthesis) but the operation which defines the chemical nature of the whole planetary surface. In summary, photosynthesis is the process by which plants use the energy of light quanta to split water H2O into hydrogen and oxygen. The oxygen bubbles off as a gas while the hydrogen reacts with CO2 to produce sugars, specifically the simple hexose sugar glucose 6CO2 + 6H2O = C6H12O6 + 6O2 (needs 2870 kJ mole-1) This reaction is plausibly believed to have started around 3.8 billion years ago (give or take a generous margin for uncertainty), and certainly changed the whole nature of our planet.
About 3800 million years ago… The earth had oceans, life, but the air was CO2/CH4 without oxygen. The oceans contained vast amounts of dissolved Iron II (blue-back). The earliest fossils are of stromatolites. Age of planet earth, billions of years 4.5 4 3.5 3 2. 5 2 1.5 1 0.5 0 Photosynthesis Oxygen builds up in air, removing Iron 2 from oceans as rust Animals Dinosaurs
Stromatolites These are our oldest fossils but can still be found (notably Shark Bay in Australia). Basically they are mounds made by photosynthetic algae which entrap sediment so accrete, and can become quite large (a metre or so) in shallow water. Humble, but we owe our existence to their slow diligent production of oxygen. Recent DNA work suggests that modern cyanophyta (blue-green algae) are relatively newly evolved, implying that the earliest stromatolites were made by unknown ancestral algae.
The key player in modern photosynthesis is an organelle called the chloroplast. We met this before under our ‘plant cells’ lecture, but need to revisit the details. You will recall that chloroplasts are derived from cyanobacteria by endosymbiosis. This bizarre process allows one organism to live inside the cell of another, gradually becoming subsumed and dependent on its larger host. (What I didn’t tell you is that this has happened hierarchically: The photosynthetic ‘dual cell’ can itself be subsumed by a larger cell, giving a chloroplast with 3 outer membranes. In the marine algae called dinoflagellates the process has occurred a 3rd time, so that their chloroplasts have 4 membranes! Almost as bizarre, many animals contain algae in their cells and rely on them partially or wholly for energy: Hydra, Cassiopeia, reef-forming corals, the giant clam Tridacna, a flatworm Convoluta.)
Chloroplasts contain many stacked light-capturing plates called thylakoids, stacked into structures discovered in Victorian times and called grana (singular granum). Grana number 10-50 in chloroplasts, and are connected together by a dense network of membranes called stroma. Granum = stack of thylakoids Contain chlorophyll Connected by lamellae The cytosol is called the stroma Ribosomes DNA Starch often stored here
Our understanding of photosynthesis derives in good part from the use of radioactive tracers. For example it is possible to supply plants with water or CO2 in which the oxygen is 18O. Heavy water H2 18O 6CO2 + 6H2 18O = C6H12O6 + 618O2 Heavy carbon dioxide C18O2 6C18O2 + 6H2O = C6H1218O6 + 6O2 What does this show? Water is the source of the O2 released during photosynthesis.
Further work (using labelled CO2 fed to algae which were given a flash of light then killed at various times afterwards) made it clear that there were many steps in the procedure, some of which needed light but some of which did not. Light reaction: needs light energy, produces ATP, a reduced electron carrier (NADPH), and H+. In order to understand this we must go into the photoelectric effect, electron transport and proton pumps. The Calvin (or Calvin-Benson) cycle, which uses ATP and NADPH CO2 to produce glucose.
The light reaction Here chlorophyll absorbs light energy and uses it to impart energy to an electron. This isn’t as odd as it might seem: light energy comes in packets or parcels called quanta (sing. quantum). Long wavelengths (red<-> infra-red) light is low energy and does little. (This is why you can put your hands under a food heater without risk of sunburn). Short wavelength (blue -> ultra violet) quanta pack more energy and can impart enough energy to a molecule to increase its energy state to a new (quantised) value. This energy state is unstable and tends to decay, with the excess energy being carried away by various means, one of which is the emission of an electron. (This is why it is the UV component of sunlight that causes the most tissue damage in sunburn).
The electromagnetic spectrum, focussing especially on the visible spectrum. High energy Low energy
The photoelectric effect One of Albert Einstein’s 3 great publications was his explanation of the photoelectric effect. When you shine light on some materials – he used the element Selenium, but plants use the molecule chlorophyll – electrons are emitted. In the case of selenium they fly off as radiation and can be measured directly (along with their energy). Chlorophyll is a bit more tricky… If you turn up the intensity of the light more electrons are emitted, but they all have the same energy. If you make the light bluer the energy of electrons increases. This was explained as light hitting the surface as bullets (quanta), with 1 quantum displacing one electron. Bluer light is equivalent to higher speed bullets.
Blue photon High E, able to displace an electron The photoelectric effect Red photon Low E, only able to warm up surface Electron e Selenium Energy of emitted electron Bluegreenred Wavelength of light Energy of photon
Chlorophyll behaves much the same way as selenium, except that the critical wavelength above which no electrons are released is c 730nm, just beyond the visible into the IR. Unlike selenium the efficiency of capture is not a simple function of wavelength but has a 2-peaked distribution: c 680nm (red) and c 425nm (blue). My Dad used to say that his house plants always died because they needed the UV that window glass filtered out. For just the once, he was wrong about this (their morbidity reflecting instead the level of care received…): plants do not need UV, indeed are damaged by it.
The chlorophyll molecules At the heart of the chlorophyll molecule lies an atom of metal held in a ring structure, much like the iron atom held in the haem group of haemoglobin. Unlike haemoglobin, chlorophyll contains magnesium. Note that there are 2 forms: Chlorophyll_a absorbs more into the UV than Chlorophyll_b. Haem ring Hydrophobic tail, anchors into phospholipid membranes
Accessory molecules (and why plants are green). If plants relied ONLY on chlorophyll they would be largely unable to use much of the visible EM spectrum. Natural selection has favoured ways to capture more energy, which is done by means of Accessory Pigments. These are other coloured chemicals, which absorb light in the visible region and pass their energy onto adjacent molecules of chlorophyll. These accessory pigments take various chemical forms – the commonest are carotenoids such as ß carotene, which helps capture longer-wavelength blue light. Together this means that plants can use energy from much of the visible spectrum. There is a dip in efficiency in the green region, for a quirk of evolution that never seems to have been explained (and probably lies in quantum physics). Why does this explain the colour of plants? (Because the colour we see is what reflects off unused.)
Antenna systems Accessory pigments are held in a tight array surrounding a chlorophyll molecule, and when they intercept a photon one of 2 things can happen: 1: Fluorescence: The energy is re-released as a photon of light. No chemical change takes place. (Ponder the wavelength of the re-emitted light: longer, same or shorter than before?) Answer: Longer – some energy will have been lost in the conversion. 2: The energy is passed along to the local reaction centre, a chlorophyll molecule. The energy is passed as an electron from molecules that receive high-energy quanta to progressively lower energy molecules, and the lowest energy energy chlorophyll molecule is the reaction centre. Its job is to act as a reducing agent.
REDOX – mnemonics, just in case you forgot… Redox state concerns the presence/absence of free electrons. It is measured in volts, but here all we need worry about is the movement of electrons. Loss of an electron = Oxidation Gain of an electron = Reduction Loss of an Electron = Oxidation Gain of an Electron = Reduction Think of a lion: LEO says GER Chemists prefer the mnemonic OILRIG. Work it out… GER
Chlorophyll donates an electron so is a reducing agent. As it happens normal chlorophyll is not much of a reducing agent, but excited chlorophyll is. Here I use * to indicate the presence of a high-energy electron. Chl* + A Chl+ + A- This redox reaction would not occur in the dark. Once the electron has been transferred to an acceptor molecule, the energy can be passed on to other molecules (always with some loss of energy). In fact there turn out to be two separate pathways for electrons to flow, making ADP -> ATP and NADP+ -> NADPH. ADP, ATP: Adenine di/tri phosphate – the unit of energy NADP – Nicotinamide adenine dinucleotide phosphate These two molecules together are the raw materials needed to make the Calvin cycle work.
Non-cyclic flow Here water is split into oxygen, H+ and electrons. It is these electrons which replace those emitted by chlorophyll and captured by NADP+ 2 different photosystems are used, called photosystem 1 and photosystem 2. They have different reaction centres and different absorbance maxima. Name abs max role Ps1 700nm uses light energy to reduce NADP+ to NADPH Ps2 680nm uses light to split water: H2O -> ½ O2 + 2H+ + 2ε-
This stage pumps protons across the thylakoid membrane for making ATP. Energy ε- ε- PS1 ε- PS2 ε- Ferrodoxin Phaephytin I 2H+ O2 H2O NADP reductase NADPH NADP+ Time This probably arose from an earlier system used by photosynthetic bacteria which oxidise hydrogen sulphide to sulphur: instead of making O2 gas they make solid S
Cyclic electron flow The non-cyclic electron flow which transfers an electron from water to NADP+ makes about equal amounts of ATP and NADPH. In fact the Calvin cycle (the CO2-fixing reaction, that takes place in the stroma) needs more ATP than NADPH, and a second mode of photosynthesis takes place in which electrons are cycled, simply making ATP. Here the high energy electron from P700 is captured by ferrodoxin and moved down a chain of cytochromes which acts as a proton pump: the energy of the electron is used to pump a proton into the thylakoid interior.
Cyclic electron flow only makes ATP (via a proton pump) Energy ε- PS1 Ferrodoxin Proton pumping chain
Proton pumps and ATP manufacture We need to explore the uses and operations of proton pumps: these are crucial to understanding both chloroplasts and mitochondria. Protons are simply H+, a hydrogen that has lost its outer electron. Well, actually no, it is a subatomic particle, which is immediately captured by any passing molecule. In fact in water when we say H+ we tend to mean H3O+, which is a molecule of water that has absorbed an extra proton, a hydroxonium ion. H2O + H+ => H3O+ Almost always when one talks of H+ one means this hydroxonium ion. Actually in this case it is indeed a proton that gets pumped across a membrane as a way of storing energy (like pump-storage scheme dams), although as soon as released into the solution it becomes H3O+.
Both in chloroplasts and in mitochondria, the only way that the protons can diffuse down the steep concentration gradient is to go through an ATP synthetase, and enzyme which performs the reaction ADP + Pi => ATP This is of course the basic unit of energy in the cell. The same operation works in the christae of mitochondria and the thylakoids of chloroplasts. The actual shape of the inner membranes are very different but more importantly the topologies are different. In mitochondria, H+ is pumped out of the innermost layer, while in chloroplasts they are concentrated inside this layer.
In a mitochondrion, H+ is pumped from the inside of the double membrane to the middle space. H+ H+ H+ H+ H+ H+ This charge gradient is maintained by several different energy-consuming enzymes (cytochrome c oxidase / reductase, NADH-Q reductase), and cannot be relieved by simple diffusion because the phospholipid bilayer is charge-impermeable, like a plastic sheet. Christae of mitochondria, H+ H+ H+ Outer membrane Inner membrane ADP + Pi ATP The protons CAN diffuse down the charge gradient (pulled by the Proton Motive Force), but only by giving up enough energy to charge up a molecule of ATP. H+ H+ H+ H+ H+ H+ H+
In a Chloroplast, H+ is pumped inside the thylakoid membrane. Otherwise the operation is exactly the same as in a mitochondrion. The pH differences can be sharp - in the light pH falls to 5 inside and rises to 8 outside - a concentration difference of how much? 1000 = antilog10(3) H+ H+ H+ H+ H+ pH5 Outer membrane Inner (thylakoid) membrane ADP + Pi ATP H+ H+ H+ H+ H+ H+
A brief insight into the advancement of science This idea, about using a proton gradient to generate ATP, is known as the chemi-osmotic hypothesis, and is an example of a maverick theory becoming standard accepted dogma. When I first learnt about chloroplasts and mitochondria (1979) we we told that the details were not fully understood, and that there was this wacky idea about acid pumps…. That’s how science develops, why science keeps changing, and why science’s focus on the operation of things gets tighter and tighter. Contrast this with religions, who are stuck with immutable words in some book written long ago, however unlikely they look…
The Calvin cycle (making sugars from CO2) Also known as the Calvin-Benson cycle. In this process high energy molecules made by light (ATP + NADPH) are used to drive the reduction of CO2. This is sometimes called the dark reaction, as light is not directly used, but the ATP etc are very short lived and in practice this cycle only happens in the light. (So don’t call it that) Our understanding of it dates to work by Calvin & Benson in the 1950s using labelled CO2 (14CO2), applied to algae in the dark, given a flash of intense light then killed in boiling ethanol a few seconds later. If the algae were killed 30s later the labelled 14C ended up in many chemicals. If they were only killed 3s after the the flash all the label was in one molecule: 3-phosphoglycerate (3PG). O 3PG H-O-C-C-C-O-P Here I use the convention that single hydrogens are omitted and P refers to a phosphate group PO43-
The process has a similarity to the citric acid cycle, by which glucose is oxidised releasing energy. In this case an initial CO2 acceptor is entered into a cyclical reaction in which a larger molecule is made, and the initial CO2 acceptor is reformed. The initial CO2 acceptor is ribulose 1,5 bisphosphate, a 5 carbon phospho-sugar. It accepts CO2 to make a 6 carbon sugar, which is then split to make 2 molecules of 3PG. This all happens in the stroma – the space outside the thylakoid membranes, into which H+ is pumped. Cleaves here rubisco C P-O-C-C-C-C-C-O-P O P-O-C-C-C-C-C-O-P CO2 + ribulose 1,5 bisphosphate O2 O H-O-C-C-C-O-P 3PG 2*
The limiting factor on the rate of photosynthesis in the light is the fixation of CO2. Specifically, the rate-limiting step is the catalysis by the enzyme Rubisco (= Ribulose bisphosphate carboxylase/oxygenase) of the uptake of CO2 by ribulose 1,5 BP. Apart from improving the efficiency of rubisco (which is presumably at an local evolutionary high point and is unlikely to improve much now), all that a plant can do to speed up the reaction is to make MORE rubisco. And they do, in fact it dominates their proteome. >20% of all the protein in plants is rubisco, making it the commonest protein in the world. This seems odd since this is an intangible globular protein – if asked most people would have guessed that animal connective proteins (collagen/keratin) would be the commonest. Rubisco Any deviation from this position causes a severe reduction in efficiency so will be selected against efficiency Configuration (AA sequence)
The 3 parts of the Calvin-Benson Cycle This cycle can be divided up into 3 parts (this is more meaningful for our visualisation of the process than it reflects reality in the plant). 1: Fixation of CO2 – catalysed by rubisco and generating 3PG. This is an acid not a sugar. 2: Conversion of PG into a sugar, in this case a simple 3 carbon sugar called glyceraldehyde 3-phosphate (confusingly abbreviated to G3P). 3: Regeneration of the CO2 acceptor, ribulose 1,5 bisphosphate.
The core of the Calvin-Benson cycle CO2 Ribulose 1,5BP ATP 3PG A series of reactions (more complex than we need to learn here today) takes P3G and, with the energy of ATP, makes ribulose. ATP, NADPH P3G
But this makes nothing – where is the output? The answer is that NOT ALL the P3G is recycled to make Ribulose. A portion is instead used in another set of reactions that make glucose, thence other complex sugars. This makes sense if we look at the numbers of carbon atoms at each stage in the cycle: Note that 6 carbons enter and 6 are withdrawn as P3G. 6* CO2 6* Ribulose 1,5BP 6 * ATP 12 * ATP + NADPH 12 *3PG 2 * C3 molecules taken away to make sugars 10 * C3 molecules enter here 12 * P3G
Rubisco and its problems In addition to capturing CO2, rubisco cleaves oxygen (adding it to make the aldehyde group in 3PG). There is in fact a direct competition between oxygen and the Calvin-Benson cycle: a curious and apparently non-adaptive reaction called photorespiration actually generates CO2 under high-oxygen levels. The factor which determines which reaction dominates is the O2:CO2 balance inside the leaf. To make sugars, leaves need high levels of CO2 and low oxygen. Some plants have evolved a special pattern of cell differentiation in their leaves to concentrate CO2 in cells using rubisco. (This is not easy as CO2 is only c. 0.04% in the atmosphere). When I learned this in 1979 CO2 was quoted as 330ppm. As I type in 2002 the official level is approaching 400ppm, and in industrial countries >400ppm is routine. This WILL affect your future.
C3 plants Most plants are C3. This means the the CO2 is captured first by incorporation into a 3 carbon molecule. What was it called again? These plants mainly have chloroplasts in the epidermal and mesophyll cells – few in the bundle sheath cells which surround the veins. Upper epidermis Mesophyll cells, with many chloroplasts (hence rubisco) Bundle sheath cells, with few chloroplasts. Sieve tube members of the vein Lower epidermis
C4 plants The trouble with being a C3 plant develops towards the end of a hot dry sunny day (when stomata are closed to conserve water) the levels of CO2 in the leaf fall very low, oxygen is high, and photosynthesis stops. (Light energy captured is fed into the photorespiration reaction instead). There is a select group of plants which have evolved to bypass this, and carry on photosynthesising after long dry periods without photorespiration cutting in. These are the C4 plants. Their special technique is to capture CO2 into a different molecule, generating a 4-carbon compound. This 4C molecule is then moved to (or stored for use by) to the rubisco-containing cells where it gives up its CO2, generating a high local concentration of the gas. This ensures that rubisco works efficiently without switching to photorespiration.
The receptor molecule C4 plants is a 3C atom called phosphoenol pyruvate (PEP), which becomes oxaloacetate via the action of an enzyme called PEP carboxylase. PEP carboxylase turns out to have advantages over rubisco: PEP carboxylase has no oxygenase activity (no losses to photorespiration), and will fix CO2 at very low concentrations. Its use allows the plant to maintain rubisco at a good CO2/O2 balance even when ambient CO2 is very low. This means that towards the end of a sunny day C4 plants will capture more energy that an equivalent C3 plant. A small point, which apparently evolved quite recently (c.12 MYBP) in response to the long-term decline in CO2. Dinosaurs breathed air 4* enriched in CO2 compared to us.
Leaf anatomy of a C4 plant Chloroplasts containing PEP carboxylase Chloroplasts containing rubisco. Upper epidermis Mesophyll cells where the chloroplasts make oxalo-acetate. Bundle sheath cells, with many chloroplasts. Also a decarboxylase which releases CO2 from oxaloacetate. Lower epidermis vein
C4 separation in time and space (Re-iteration of what you already know:) The possession of C4 chloroplasts allows plants to capture CO2 efficiently even at very low levels and store the gas as oxaloacetate. This chemical is ‘equivalent’ to CO2 since it can readily be decarboxylated to release CO2. Some tropical grasses (maize, sorghum, crabgrass) use this trick to carry on photosynthesising down to very low internal CO2 concentrations. Here the rubisco chloroplasts are separated from the PEP carboxylase chloroplasts in space, being physically segregated in the bundle sheath cells. (This bit is new:) Another group of plants use the same trick to ensure that they never need open their stomata during daylight. Here the rubisco and PEP are separated in time.
CAM plants CAM plants are adapted to dry conditions. This has evolved several times, and is named after the Crassulas (thick-leaved succulent dicots preferring very dry conditions.) CAM = Crassulacean acid metabolism. In the UK we have a few native crassulacea - the ? stonecrops Sedum! Houseleeks (Sempervivum) are another in this family. The unrelated cacti also have CAM, as do pineapples. Here the oxaloacetate path is used at night to open the stomata. The point is that the stomata are never opened by day, but open instead at night when conditions are cooler.
In CAM plants The C4 compound oxaloacetate is stored as another C4 compound malic acid, which is then decarboxylated during the day by the normal photosynthetic processes. Night Day Night Malic acid C4 using stored ATP etc Photosynthesis makes 3PG using rubisco as normal, also ATP. CO2 Stomata shut Stomata shut Stomata shut