Enzyme Kinetics
Enzyme Kinetics. E. S. P. k. k. 2. 1. <->. ->. E. +. S. ES. P. +. E. k. 1. -. Enzyme Kinetics I. An enzyme-catalyzed reaction of substrate S to product P, can be written


Enzyme Kinetics
E N D
Presentation Transcript
E S P k k 2 1 <-> -> E + S ES P + E k 1 - Enzyme Kinetics I An enzyme-catalyzed reaction of substrate S to product P, can be written Actually, the enzyme and substrate must combine and E recycled after the reaction is finished, just like any catalyst. Because the enzyme actually binds the substrate the reaction can be written as: The simplest reaction is a single substrate going to a single product.
Rate or velocity of the reaction depends on the formation of the ES • The P -> ES is ignored • The equilibrium constant Keq is based on the idea that the reaction is limited to the formation of the ES complex and that only K1 and K-1 are involved because the thermodynamics of the reversal of K2 cause it to be minimal How fast an enzyme catalyzes a reaction is it's rate. The rate of the reaction is in the number of moles of product produced per second k 1 Keq = K - 1 d[P] rate (v) = = k [ES] 2 dt
The relationship between the concentration of a substrate and the rate of an enzymatic reaction is described by looking at the concentration of S and v • When the reaction is first order - the rate is dependent on [S] • When the reaction is zero order, there is no relationship between v and S • A second order is between 1st and 0 order, where the relationship between V and [S] is not proportional to [S] Initial Velocity (Vi or V) [ Substrate]
To study enzymes, first order kinetics must be followed! • Think of the graph of [S] vs. v in this way: • The velocity increases as the substrate concentration is increased up to a point where the enzyme is "saturated" with substrate. • At this point the rate of the reaction (v) reaches a maximal value and is unaffected by further increases in substrate because all of the enzyme active site is bound to substrate
For the most part enzyme reactions are treated as if there is only one substrate and one product. If there are two substrates, one of them is held at a high concentration (0 order) and the other substrate is studied at a lower concentration so that for that substrate, it is a first order reaction. This leads us to the M and M equation.
Conditions for Michaelis -Menten Two assumptions must be met for the Michaelis-Menten equation • Equilibrium -the association and dissociation of the substrate and enzyme is assumed to be a rapid equilibrium and Ks is the enzyme:substrate dissociation constant.
Conditions for Michaelis -Menten Two assumptions must be met for the Michaelis-Menten equation • Steady state - the enzyme substrate complex ES is at a constant value. That is the ES is formed as fast as the enzyme releases the product. For this to happen the concentration of substrate has to be much higher than the enzyme concentration. That is why we only study the initial velocity. Later in the reaction the substrate concentration is relatively lower and the rate of product starts to be limited by diffusion and not the mechanism of the enzyme.
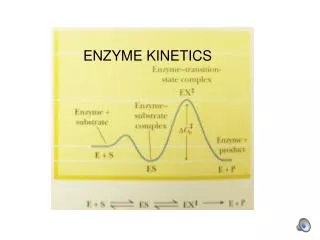
Michaelis-Menten Enzyme kinetics • Don't for get the two assumptions - They both lead to the same equation, the michaelis-menten equation. • What is this awe inspiring equation? The Michaelis-Menten kinetic model explains several aspects of the behavior of many enzymes. Each enzyme has a Km value that is characteristic of that enzyme under certain conditions.
Graphical model of the representation of the M&M eq. • Reaction velocity (V) vs concentration of substrate [S] • - as [S] increases, velocity increases and eventually levels off = V max • 1st order vs zero order rates of reaction - back to the two assumptions • There are two important values for each enzyme that are described by the M&M equation; V max and Km (Michaelis-Menten constant) • Graphically, these are shown as 1/2 V max = Km can not reach real V max so....
k k 2 1 <-> -> E + S ES P + E k 1 - K + K - 1 2 K = m K 1 • Mathematical model of the representation of the M&M eq. - For the reaction: 1) The Michaelis constant Km is: Think of what this means in terms of the equilibrium. Large vs. a small Km
2) When investigating the initial rate (Vo) the Michaelis-Menten equation is: Graphical representation is a hyperbola. Think of the difference between O2 binding of myoglobin and hemoglobin. • When [S] << Km, the velocity is dependent on [S] • When [S] >> Km, the initial velocity is independent of [S] • When [S] = Km, then Vo = 1/2 V max Prove this mathematically and graphicaly. V [S ] m a x V o = [ S ] + K m
Km is a measure of the affinity of the enzyme for it's substrate and also informs about the rate of a reaction. The binding constant is appoximated by Km • Rules for using the M&M equation: • The reaction must be first order and [S] >> E (two assumptions)
Turnover Number - kcat - the direct measure of the catalytic production of product. The larger the kcat is, the more rapid the catalytic events at the enzyme's active site must be. The number of times a binding and reaction event "turns over" • When the [S] << Km so that most of the enzyme is in the free state [E]t = [E]free then V = kcat / Km [E][S] • This is a second order rate constant between the substrate and the free enzyme. This is a good measure of efficiency and specificity. • When the kcat/Km is near very high, the fastest the enzyme can catalyze a reaction is the diffusion rate of a molecule! 108 - 109 / M . sec
Lineweaver-Burk (double reciprocal plot) • Vmax and Km are not likely to be determined by increasing [S] • Instead the [S] vs. Vo data are transformed to a plot of their reciprocal of each value. • 1/[S] vs. 1/Vo
So What? • Km - relates to affinity ; Vmax relates to efficiency • Km tell how much substrate to use in an assay • If more than one enzyme share the same substrate, KM also will determine how to decide which pathway the substrate will take Vmax tells about pathways • Rate limiting enzyme in pathway • Km andVmax can be used to determine effectiveness of inhibitors and activators for enzyme studies and clinical applications
Enzyme inhibitors Competitive inhibition • Inhibitor is similar to substrate and both bind to or near active site. compete’ for binding • inhibitor is unreactive - EI state • Lineweaver Burke intersect at the Y axis
Enzyme inhibitors Noncompetitive inhibitor • inhibitor binds distal to active site • effects enzyme rate not affinity • binds E in E S or E • Reversible • Lineweaver Burke intersect at the Y axis
Mixed Inhibition • Inhibitor binds to enzyme site that involves both S binding and catalysis • binds E in E S or E • Forget the alpha business
Enzyme inhibitors Uncompetitive inhibitor • binds covalently in the transition state • suicide inhibitor • binds to the ES complex • lowers affinity and velocity • lineweaver Burke plots are parallel
Penicillin as a suicide substrate • - suicide substrates are often un competitive inhibitors that decrease the energy of the transition state and allow the ES to have lower energy that that of the EP. • Bacterial cell wall - extensive cross linking of sugars and peptides • Penicillin (and ampicillin) have a highly reactive ß lactam ring which makes a peptide bond very reactive.
Penicillin as a suicide substrate • Penicillin mimics the peptide alanine residues and forms a low energy intermediate by covalently reacting with a serine • In molecular biology, we use this as a tool. Ampicillin will stop E. coli growth. Bacteria that have a gene (plasmid) inserted into the bacteria have ß lactamase. An enzyme that hydrolyses the reactive peptide bond found in amicillin and penicillin
Allosteric Regulation • An organism must be able to regulate the catalytic activities of its component enzymes • coordinate many metabolic processes • Respond to changes in the environment • Growth and differentiation • Both Inhibitors and affectors
Allosteric Regulation • do not follow Michaelis-Menten kinetics - instead use a hill plot for both + and – effects • similar to O2 dissociation of hemoglobin
Allosteric Regulation • Two ways: • Control enzyme availability • Synthesis of enzyme • Degeneration • Control enzyme activity • Alterations which affect the substrate binding affinity • Turn over number
Allosteric Regulation • Can cause large changes in enzymatic activity • Regulated by covalent modifications • Usually Phosphorylation and de-Phosphorylation of specific Ser and Tyr residues.
Phosphorylation • Phosphorylation is the addition of a phosphate (PO4) group to a protein or other organic molecule. • kinases (phosphorylation) and phosphatases (dephosphorylation) are involved in this process. Many enzymes and receptors are switched "on" or "off" by phosphorylation and dephosphorylation. • Reversible phosphorylation results in a conformational change in the structure in many enzymes and receptors, causing them to become activated or deactivated. Phosphorylation usually occurs on serine, threonine, and tyrosine residues in eukaryotic proteins
Phosphorylation regulates phosphenol-pyruvate (PEP) carboxylase • CAM and C4 plants require a separation of the initial carboxylation from the following de-carboxylation • Diuranalregulation is used • IN CAM PLANTS:- • Phosphorylation of the serine residue of phosphenol-pyruvate (PEP) carboxylase (Ser-OP) yields a form of the enzyme which is active at night • This is relatively insensitive to malic acid
Photophorylation regulates phosphenol-pyruvate (PEP)carboxylase • During the day: • De-Phosphorylation of the serine (ser-OH) gives a form of the enzyme which is inhibited by malic acid • THIS IS THE OPPOSITE WAY AROUND FOR C4 PLANTS!
Phosphorylation • The addition of a phosphate (PO4) molecule to a polar R group of an amino acid residue can turn a hydrophobic portion of a protein into a polar and extremely hydrophilic portion of molecule. • In this way it can introduce a conformational change in the structure of the protein via interaction with other hydrophobic and hydrophilic residues in the protein • Examples: • Phosphorylation of the cytosolic components of NADPH oxidase, plays an important role in the regulation of protein-protein interactions in the enzyme • Phosphorylation of the enzyme GSK-3 by AKT (Protein kinase B) as part of the insulin signaling pathway
Phosphorylation • There are thousands of distinct phosphorylation sites in a given cell since: • There are thousands of different kinds of proteins in any particular cell (such as a lymphocyte). • It is estimated that 1/10th to 1/2 of proteins are phosphorylated (in some cellular state). • Phosphorylation often occurs on multiple distinct sites on a given protein.
Bisubstrate Reactions • So far: • Simple, single-substrate reactions that obey the Michaelis-Menten model • However, approx 60% of known biochemical reactions involve two substrates and yield two products • Either: • transfer reactions – moving a functional group from one substrate to the other • Oxidation/reduction reaction between substrates
Bisubstrate Reactions • Sequential reactions • All substrates must combine with the enzyme before the reaction can occur and products are released • A - leading substrate • B – following substrate • P – 1st product leaving enzyme • Q – 2nd product leaving enzyme • ie NAD+ and NADH reactions involving dehydrogenases
Bisubstrate Reactions • Ping-pong reactions • Group transfer reactions in which one or more products are released before all substrates have been added. • Two stage reaction: • A functional group from 1st sub (A) is transferred to the 1st product (P) forming a stable enzyme (F) –The Ping • The functional group is displaced from the enzyme by the 2nd substrate (B) to yield 2nd product (Q), regenerating the original form of the enzyme (E) – The Pong • iemany reactions involving Trypsin