Download

1 / 87
930 likes | 1.35k Vues
Cytokinin 細胞分裂素. 生科四 林暐哲. Cytokinins 對植物的影響. Cell division Delay leaf senescence Promote nutrient mobilization Modify apical dominance The formation and activity of shoot apical meristem Floral development. Cytokinin mediate light-regulate development. Break bud dormancy

E N D
Cytokinin細胞分裂素 生科四 林暐哲
Cytokinins 對植物的影響 • Cell division • Delayleaf senescence • Promote nutrient mobilization • Modify apical dominance • The formation and activity ofshoot apical meristem • Floral development
Cytokinin mediatelight-regulatedevelopment • Breakbud dormancy • Stimulateseed gemination • Chloroplast differentiation • Development ofautotrophic metabolism • Leaf and cotyledon expansion
細胞分裂and 植物的生長 植物由primaryandsecondarymeristem(分生組織)生成新細胞 新細胞經過 enlargement and differentiation 之後,一般不再進行cell division (類似動物細胞)
但是幾乎所有類型的已分化的植物細胞(保留nucleus) 能夠再次分裂,例如植物在進行 wound healing(癒傷)和形成 leaf abscission(葉離層) http://botit.botany.wisc.edu/
differentiated plant cell can resume division • 在Cortex(皮層)及 Phloem(韌皮部)的成熟細胞重新分裂以形成secondary meristems vascular cambium(維管束形成層) cork cambium(木栓形成層)
在leaf petiole(葉柄)基部abscission zone (離層區)的成熟parenchyma cells(薄壁細胞),在有絲分裂後一段時間再度分裂,而形成較弱的細胞壁,進而使abscission產生,葉因而容易脫落
植物Wound(損傷)會誘使在損傷部位的細胞重新分裂,稱為wound-induce mitotic,甚至連高度專一的phloem fibers(韌皮纖維)及 guard cells(保衛細胞)都會再度分裂。 Wound-induce mitotic activity通常會self-limiting,表示這些細胞在分裂數次後會停止分裂並進行分化
Philip White在1930年代指出tomaoto roots能無限期的培養在一種含有簡單營養的medium中,此Medium包含蔗糖、礦物鹽類及少量vitamines,而未添加任何hormones • 但分離出的stem tissue卻無法在此medium中生長,即使加進Auxin之後仍然只有些微的生長。而auxin-induced生長也只是源於細胞enlargement卻沒有細胞分裂。
後續的研究發現,幾乎所有植物的shoot tissues都無法在medium之中培養,除非這些組織偶然長出了root,shoot才會繼續生長,但此組織最終將長成一個完整的植物體 • 由此說明shoot與root的分生組織,對於細胞分裂的調控有某些差異
Callus tissue • 若將形成不斷分裂的crown gall的植物,以42℃加熱殺死細菌,植物仍會存活而此crown gall中的tissue將會持續生長成不含細菌(bacteria-free)的tumor • 將這個tumor用medium培養,增殖的細胞將形成不規則的組織,稱為Callus tissue • Callus tissue證實了來自shoot的組織可以被培養,而這些曾與細菌接觸過的細胞會製造出促進細胞分裂的物質
Cytokinin的發現、鑑定與性質 • Caplin and Steward在1948年將Philip White的medium加入auxin和10-20%的椰奶,發現能使許多種類植物的成熟已分化的細胞恢復分裂而形成callus tissue • 多年後的研究證實椰奶中含有 cytokinin zeatin
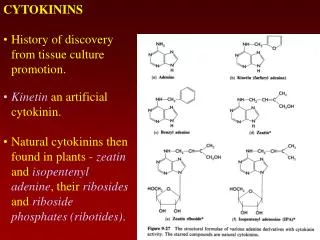
Kinetin • Folke Skoog在1940-1950年代對tobacco pith tissue增殖的實驗中發現核酸中的adenine對促進細胞分裂有些微的效果,而經過autoclave(高壓滅菌釜)處理的鯡魚精子有強大的促細胞分裂效果 • Millery在1955年將此物質由高壓滅菌處理過的DNA中分離出來並命名為Kinetin
Kinetin • Kinetin是adenine的衍生物, 6-furfurylaminopurine
Zeatin • 1963年Letham由未成熟的玉米種子胚乳中萃取出一種生物活性類似於kinetin的物質,trans-6-(4-hydroxy-3-methylbut-2-enylamino)purine,命名為zeatin • Zeatin與kinetin結構相似,都具有adenine
Zeatin • Zeatin 的側鏈上帶有雙鍵,因此有trans與cis兩種型態,在高等植物體中同時含有這兩種,並可藉zeatin isomerase互變 • trans is more active,但cis態也具重要角色,例如玉米cytokinin receptor會同時與兩種型態結合,特定受cis態影響的酵素也已被發現
具有cytokinin活性的合成分子 • 在與auxin同時存在時能夠促進callus tissue進行細胞分裂 • 促使callus生成芽或根 • 延緩葉的老化 • 促進雙子葉植物的子葉擴張
Cytokinins有游離態與結合態 • Cytokinins以游離分子存在於植物體,最多的是zeatin,而dihydrozeatin(DZ)與isopentenyl adenine(iP)也常被發現存在高等植物,許多這三種Cytokinins的衍生物也從植物萃取液中被鑑定
部份植物的tRNA具有 cis-zeatin出現在過度修飾的鹼基上
Active Cytokinin • 阿拉伯芥Cytokinin receptor CRE1是與free form的trans-zeatin結合,而不是其他riboside或ribotide的衍生物,DZ與iP也是相似的情況
微生物也能製造cytokinin • 許多與植物相關的微生物如細菌、真菌能製造大量的cytokinin,包含trans-zeatin、iP、cis-zeatin及其ribosides • 感染這些微生物的植物體會生成特殊的構造如菌根、 crown gall(A.tumefaciens) witches’broom (Corynebacterium fascian)
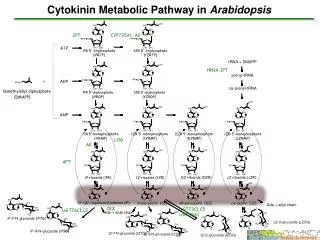
Cytokinins的合成、代謝、運輸 • Zeatin與iP的側鏈是由isoprene衍生而來 • 大分子如橡膠及胡蘿蔔素是由isoprene聚合而來 • Dimethylallyl Diphosphate (DMAPP)是植物利用提供isoprene unit • 細菌利用1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate (HMBDP)
Crown gall cells具有合成cytokinin的基因 • 造成crown gall的細菌agrobacterium tumefaciens含有一段大型質體稱為Ti-plasmid,當植物感染此細菌時 Ti-plasmid會鑲入植物染色體DNA內進行轉錄 • Ti-plasmid中含有一段T-DNA含有合成trans-zeatin和Auxin的基因
T-DNA • 使變為crown gall的cell製造Opine 作為氮源利用 • Ipt gene → isopentenyl transferase(IPT) • Ipt gene與兩個合成Auxin的gene稱為phyto-oncogene(植物致癌基因) • 因為T-DNA的promoters為eukaryotic,所以只有在感染植物宿主時才會表現
Cytochrome P450 monooxygenase iPMP/iPDP/iPTP → ZMP/ZDP/ZTP
Cytokinin的運輸調節 • 根尖分生組織是植物主要合成cytokinin的場所,合成的cytolinin由木質部伴隨著水及礦物質向上運輸到莖部 • 木質液所含的cytolinin大部分是zeatin riboside,當被運送到葉後,會轉為游離態或glucoside • zeatin riboside的調節主要來自莖的訊息
Beveridge 在1997年將wild type的pea與mutant rms4做嫁接實驗,此rms4的木質液zeatin riboside含量只有正常的1/40 rms4 shoot + wild type root low [cytokinin] level wild type shoot + rms4 root high [cytokinin] level 由此證實木質液cytokinin運輸的調控主要來自莖部的訊息
其他合成cytokinin的部位 • 阿拉伯芥的7個IPT genes有各自對應的作用部位,除了根尖外,還包括Phloem、leaf axil、ovules、immature seeds、columella root cap、young花的上部、fruit abscission zone • 有些IPT genes受cytokinin的down-regulate 另一些IPT genes則受Auxin的 up-regulate
Cytokinin的代謝調控 • Cytokinin會被cytokinin oxydase切割成側鏈與adenine(只有O-glucoside形式與DZ不會被切割) ,以減少cytokinin的效果 • Cytokinin也會以在第3,7,9號氮上結合一個glucose殘基的方式來使其失去活性。 • O-glucoside應該是一種儲存Cytokinin的形式,能被glucosidase轉變回具活性的形式,此現象與休眠種子萌芽有關
Cytokinins 報告者:江禮憲 LINCOLN TAIZ‧EDUARDO ZEIGER
Cytokinins regulate cell division in shoots and roots • Several evidence • Shoot apical meristem (Werner et al. 2001) - This finding strongly supports the notion that endogenous cytokinins regulate cell division
Cytokinins regulate cell division in shoots and roots • Cytokinin oxidase enhancement of root growth (Werner et al. 2001) • Root apical meristem (Werner et al. 2001)
Cytokinins regulate cell division in shoots and roots • A small reduction in cytokinin levels would increase root growth (data courtesy of F. J. Ferreira and J. J. Kieber.)
Cytokinins regulate cell division in shoots and roots • Triple-receptor mutants (ahk2/ahk3/cre1) (Nishimura et al. 2004.) • Mutation in maize alters leaf phyllotaxy and the size lf the shoot apical meristem (abphyl1) (Giulini et al. 2004.)