Download
1 / 39
420 likes | 800 Vues
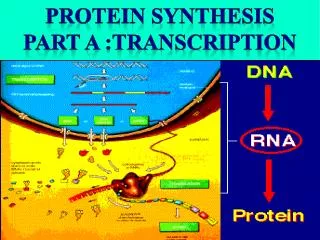
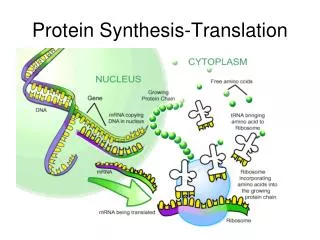
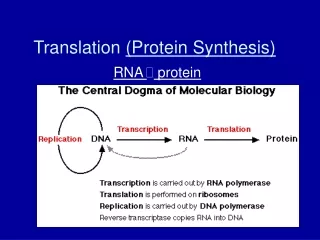
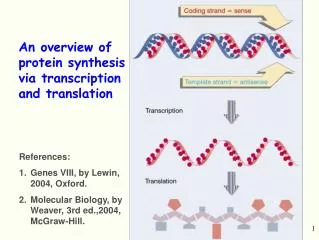
An overview of protein synthesis via transcription and translation. References: Genes VIII, by Lewin, 2004, Oxford. Molecular Biology, by Weaver, 3rd ed.,2004, McGraw-Hill. P romoter. +1. Terminator. -35. -10. Transcription (RNA polymerase). Ribosome-binding site. ORF. mRNA.

E N D
An overview of protein synthesis via transcription and translation • References: • Genes VIII, by Lewin, 2004, Oxford. • Molecular Biology, by Weaver, 3rd ed.,2004, McGraw-Hill.
Promoter +1 Terminator -35 -10 Transcription (RNA polymerase) Ribosome-binding site ORF mRNA Start codon Stop codon Translation (Ribosome) Protein Prokaryotic Gene Expression
Transcription in Prokaryotic Cells RNA polymerase DNA template Coding strand Template strand promoter; terminator RNA pol RNA pol
Stages of transcription Template recognition –Initiation – Elongation - Termination 3’ 5’
RNA pol in eubacteria core:a2bb’ holoenzyme: core + s factor s factor is separated from the core when holoenzyme is subjected to an anion exchange (e.g. phosphocellulose) column
RNA DNA Yeast RNA polymerase
Functions of s factor Effect of s factor Promoter recognition. Promote tight binding of holoenzyme to the promoter. Loosening non-specific interaction between RNA pol and template. Stimulates transcription initiation.
converts a loosely bound RNA pol in a closed complex to the tightly bound pol in the open promoter complexes. Supercoiled DNA is a better template for transcription, because it requires less free energy for the initial melting of DNA. RNA pol-promoter binding
1. Forming the closed promoter complex Template recognition 2. Forming the open promoter complex 3. Abortive initiation Initiation 4. Promoter clearance
How RNA polymerase gets to the promoter? Sliding along DNA does not occur
s factor can be reused -Rifampicin Rif R +Rifampicin Rif S Sigma cycle
RNA Pol-Promoter Interaction DNA region covered by holoenzyme is from -55 to +20; that covered by core enzyme after loss of s is from -35 to +20.
RNA Pol-Promoter Interaction Methylation Interference Assay Bases on either the template or the non-template strand that are more methylated in the filtrate than in the filter-bound DNA are presumably important in polymerase binding to the promoter.
RNA pol-promoter contact -9 to +3
-10 +1 -35 18 bp lac trp lPL recA tacI TTTACA TTGACA TTGACA TTGATA TTGACA TATGTT TTAACT GATACT TATAAT TATGTT 17 bp 17 bp 17 bp 9 bp 7 bp trp lac 18 bp TTGACA Consensus TATAAT (5-9 bp) (16-19 bp) >90% of transcription start point is a purine Features of bacterial promoters
How many kinds of s factors are there in a bacterial cell? What is the structure of s factor?
Structure of s factors Primary s factors (e.g. s70 of E. coli; s43 of B. subtilis) Alternative s factorsTranscription of specialized genes (e.g. s54 of E. coli) Free s cannot bind to the promoter (The N-terminal region suppresses the DNA-binding region). Only when it is bound with the core, upon which its conformation changes, can s binds the promoter.
245 aa deletion s70 Region 1.Present only in primary s. The 245 aa existing in s70, but not in s43, may be involved in loosening binding between RNA pol and non-promoter regions. Region 2.Most highly conserved. 2.1 and 2.2: hydrophobic; binding to pol core. 2.3: involved in DNA melting. 2.4: a-helix; recognition of -10 box. Region 3.Helix-turn helix DNA-binding domain. Region 4. 4.2: helix-turn-helix loop; binding to -35 box.
Primary vs. alternative s factors in E. coli Different s factors recognize promoters with different consensus sequences • s54 is different from other s factors in: • The “-35 box” is located 6 bp upstream of the “-10 box”; • Sites that are rather distant from the promoter influence its activity (recognized by an enhancer-binding protein); • The free form can bind to DNA.
Sigma-switching model Temporal control of transcription of B. subtilis phage SPO1
Genetic evidence Isolation of mutants that are unable to do transcription switch. Biochemical evidence Composition analysis of the RNA pol isolated from different stages. Other examples : Control of transcription during sporulation in B. subtilis Regulation of glutamine synthetase gene (s54) Regulation of the E. coli heat shock genes (s32) Stress-resistance genes turned on in the stationary phase (ss)
Elongation Functions of RNA pol core • To unwind and rewind DNA • To hold the separated strand of DNA and RNA • To catalyze the addition of ribonucleotides to the growing RNA chain • To adjust the difficulties in processing by cleaving the RNA product and restarting RNA synthesis (with the assistance of some accessory factors, e.g., GreA and GreB in E. coli)
Function of a-subunit Core enzyme assembly; Promoter recognition; Interaction with some regulators. Role of a subunit in UP element recognition: 1. Addition of UP to the core promoter increases in vitro transcription by RNA pol alone. 2. The 94 C-terminal aa are required for UP recognition.
UP element: an AT-rich sequence which stimulates transcription of the rrnB gene by a factor of 30. Fis sites: binding sites for Fis, a transcriptional activator.
Function of b and b’-subunit b-subunit Phosphodiester bond formation. (Confers both the rifampicin- and streptolydigin-resistance) Stabilizing RNA pol-DNA complex during elongation. Forms both the salt-sensitive and salt-resistant contact with the DNA template. b’-subunit Most basic subunit. Strongest DNA-binding activity. Forms salt-resistant contact with the DNA template.
Topology of elongation The strain of unwinding is relaxed by the topoisomerases.
Termination Mechanism r-independent termination (intrinsic terminators) Requires: a hairpin loop a string of Ts following the hairpin.
Intrinsic terminators Stem of hairpin: G-C-rich; 7-20 bp Loop: 5 bp or up Distance between hairpin and U-run: 7-9 bp
r-dependent termination Half of E. coli terminators; most are found in phage genomes Requires a hairpin loop and r factoracts as a homohexamer each subunit contains an RNA-binding domain an ATPase domain. an RNA helicase (separates RNA-DNA hybrid)
r-dependent terminator 50-90 bases long C-rich/G poor
Polar effect on transcription of the downstream genes caused by a nonsense mutation
Negative control: Control of prokaryotic transcription Repressor Inducible genes v.s. Constitutive genes Positive control: Activator
Cofactors of the regulators: Repressor Inducer Corepressor Activator Inducer Common features of the cofactors: Highly specific Not necessarily interact with the target enzyme Gratuitous inducers (e.g., IPTG) Allosteric control of the regulator Other means of activation of regulators: Phosphorylation Oxidation Other positive control mechanisms: Substitution of s factors Antitermination
Operon A group of contiguous, coordinately controlled genes The lac operon The first operon discovered (Jacob and Monod, 1961) Polycistronic mRNA 4