Download
1 / 1
10 likes | 104 Vues
709.27. Introduction * The serotonin (5-HT) neurotransmitter system is thought to play a key role in the regulation of mood, anxiety states and motor activity [1].

E N D
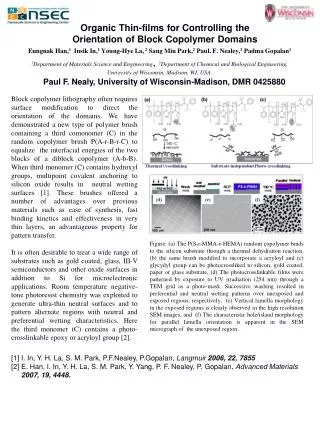
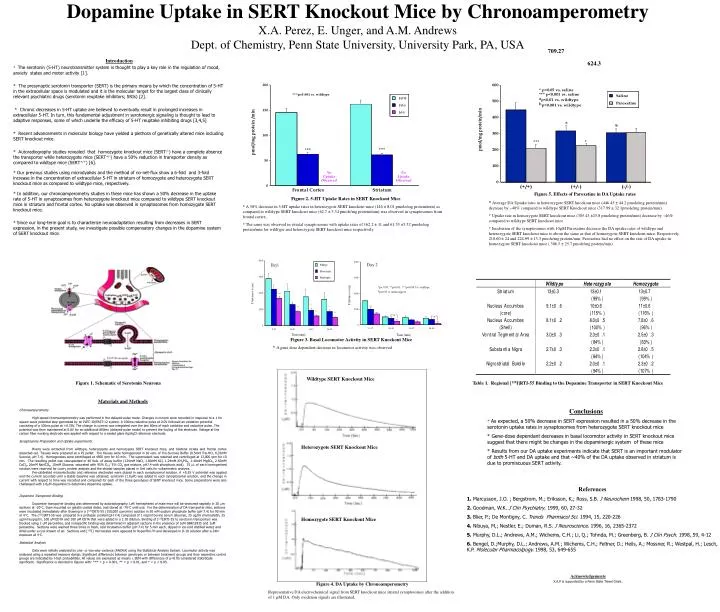
709.27 Introduction * The serotonin (5-HT) neurotransmitter system is thought to play a key role in the regulation of mood, anxiety states and motor activity [1]. * The presynaptic serotonin transporter (SERT) is the primary means by which the concentration of 5-HT in the extracellular space is modulated and it is the molecular target for the largest class of clinically relevant psychiatric drugs (serotonin reuptake inhibitors; SRIs) [2]. * Chronic decreases in 5-HT uptake are believed to eventually result in prolonged increases in extracellular 5-HT. In turn, this fundamental adjustment in serotonergic signaling is thought to lead to adaptive responses, some of which underlie the efficacy of 5-HT reuptake inhibiting drugs [3,4,5] * Recent advancements in molecular biology have yielded a plethora of genetically altered mice including SERT knockout mice. * Autoradiography studies revealed that homozygote knockout mice (SERT-/-) have a complete absence the transporter while heterozygote mice (SERT+/-) have a 50% reduction in transporter density as compared to wildtype mice (SERT+/+) [6]. * Our previous studies using microdyalisis and the method of no-net-flux show a 6-fold and 3-fold increase in the concentration of extracellular 5-HT in striatum of homozygote and heterozygote SERT knockout mice as compared to wildtype mice, respectively. * In addition, our chronoamperometry studies in these mice has shown a 50% decrease in the uptake rate of 5-HT in synaptosomes from heterozygote knockout mice compared to wildtype SERT knockout mice in striatum and frontal cortex. No uptake was observed in synaptosomes from homozygote SERT knockout mice. * Since our long-term goal is to characterize neuroadaptation resulting from decreases in SERT expression, in the present study, we investigate possible compensatory changes in the dopamine system of SERT knockout mice. Dopamine Uptake in SERT Knockout Mice by ChronoamperometryX.A. Perez, E. Unger, and A.M. AndrewsDept. of Chemistry, Penn State University, University Park, PA, USA624.3 Figure 5. Effects of Paroxetine in DA Uptake rates * Average DA Uptake rates in heterozygote SERT knockout mice (446.45 ± 44.2 pmole/mg protein/min) decrease by ~40% compared to wildtype SERT Knockout mice (317.99 ± 32.1pmole/mg protein/min). * Uptake rate in homozygote SERT knockout mice (305.43 ±25.8 pmole/mg protein/min) decrease by ~46% compared to wildtype SERT knockout mice * Incubation of the synaptosomes with 10M Paroxetine decrease the DA uptake rates of wildtype and heterozygote SERT knockout mice to about the same as that of homozygote SERT knockout mice. Respectively, 210.60 ± 24 and 224.99 ± 13.3 pmole/mg protein/min. Paroxetine had no effect on the rate of DA uptake in homozygote SERT knockout mice ( 306.5 ± 25.7 pmole/mg protein/min) Figure 2. 5-HT Uptake Rates in SERT Knockout Mice * A 50% decrease in 5-HT uptake rates in heterozygote SERT knockout mice (146 ± 8.01 pmole/mg protein/min) as compared to wildtype SERT knockout mice (62.7 ± 3.34 pmole/mg protein/min) was observed in synaptosomes from frontal cortex. * The same was observed in striatal synaptosomes with uptake rates of 162.2 ± 11 and 61.53 ±5.52 pmole/mg protein/min for wildtype and heterozygote SERT knockout mice respectively. Acknowledgements X.A.P is supported by a Penn State Travel Grant. Figure 3. Basal Locomotor Activity in SERT Knockout Mice * A gene dose dependent decrease in locomotor activity was observed Wildtype SERT Knockout Mice Figure 1. Schematic of Serotonin Neurona Table 1. Regional [125I]RTI-55 Binding to the Dopamine Transporter in SERT Knockout Mice Materials and Methods Chronoamperometry High-speed chronoamperometry was performed in the delayed-pulse mode. Changes in current were recorded in response to a 1 Hz square wave potential step generated by an IVEC-10/FAST-12 system. A 100ms reductive pulse at 0.0V followed an oxidation potential consisting of a 100ms pulse at +0.55V. The change in current was integrated over the last 80ms of each oxidative and reductive pulse. The potential was then maintained at 0.0V for an additional 800ms (delayed-pulse mode) to prevent the fouling of the electrode. Voltage at the carbon fiber working electrode was applied with respect to a sealed glass Ag/AgCl reference electrode. Synaptosome Preparation and Uptake experiments. Brains were extracted from wildtype, heterozygote and homozygote SERT knockout mice, and bilateral striata and frontal cortex dissected out. Tissues were prepared as a P2 pellet. The tissues were homogenized in 10 vols. of Tris-Sucrose Buffer (0.5mM Tris-HCl, 0,32mM Sucrose, pH 7.4). Homogenates were centrifuged at 4500 rpm for 10 min. The supernatant was retained and centrifuged at 13,000 rpm for 10 min. The resulting pellet was resuspended in 40 Vols. of assay buffer (124mM NaCl, 1.80mM KCl, 1.24mM KH2PO4, 1.40mM MgSO4, 2.50mM CaCl2, 26mM NaHCO3, 10mM Glucose, saturated with 95% O2 / 5% CO2 gas mixture, pH7.4 with phosphoric acid). 25 L of each homogenized solution were reserved for Lowry protein analysis and the striatal samples placed in 3ml vials for voltammetric analysis. Pre-calibrated microelectrodes and reference electrodes were placed in each synaptosomal solution. A +0.55 V potential was applied and the current recorded until a stable baseline was obtained. serotonin (1.0M) was added to each synaptosomal solution, and the change in current with respect to time was recorded and compared for each of the three genotypes of SERT knockout mice. Some preparations were also challenged with 1.0M dopamine to determine dopamine uptake. Dopamine Transporter Binding Dopamine transporter binding was determined by autoradiography. Left hemispheres of male mice will be sectioned sagitally in 20 m sections at -20C, thaw mounted on gelatin coated slides, and stored at -70C until use. For the determination of DA transporter sites, sections were incubated immediately after thawing in a [125I]RTI-55 (150,000 cpm/min) solution in 50 mM sodium phosphate buffer (pH 7.4) for 90 min at 4C. The [125I]RTI-55 was prepared in a protease cocktail (pH 7.4) composed of 1 mg/ml bovine serum albumen, 25 ug/ml chymostatin, 25 ug/ml leupeptin, 100 uM EDTA and 100 uM EGTAthat were added in a 1:10 dilution. Binding of [125I]RTI-55 to serotonin transporters was blocked using 1 uM paroxetine, and nonspecific binding was determined in adjacent sections in the presence of 1uM GBR12935 and 1uM paroxetine. Sections were washed three times in fresh, cold incubation buffer (pH 7.4) for 5 min each, dipped in ice-cold distilled water, and dried under a cool stream of air. Sections and [125I] microscales were apposed to Hyperfilm-3H and developed in D-19 solution after a 24hr exposure at 4C. Statistical Analysis Data were initially analyzed by one- or two-way variance (ANOVA) using the Statistical Analysis System. Locomotor activity was analyzed using a repeated measure design. Significant differences between genotypes or between treatment groups and their respective control groups are indicated by t-test probabilities. All values are expressed as means SEM with differences of p<0.05 considered statistically significant. Significance is denoted in figures with: *** = p < 0.001, ** = p < 0.01, and * = p < 0.05. Conclusions * As expected, a 50% decrease in SERT expression resulted in a 50% decrease in the serotonin uptake rates in synaptosomes from heterozygote SERT knockout mice * Gene-dose dependant decreases in basal locomotor activity in SERT knockout mice suggest that there might be changes in the dopaminergic system of these mice * Results from our DA uptake experiments indicate that SERT is an important modulator of both 5-HT and DA uptake and that ~40% of the DA uptake observed in striatum is due to promiscuous SERT activity. Heterozygote SERT Knockout Mice References 1. Marcusson, J.O. ; Bergstrom, M.; Eriksson, K,; Ross, S.B. J Neurochem 1998, 50, 1783-1790 2. Goodman, W.K. J Clin Psychiatry. 1999, 60, 27-32 3. Blier, P.; De Montigny, C. Trends Pharmacol Sci. 1994, 15, 220-226 4. Nibuya, M,; Nestler, E.; Duman, R.S. J Neuroscience. 1996, 16, 2365-2372 5. Murphy, D.L.; Andrews, A.M.; Wichems, C.H.; Li, Q.; Tohnda, M.; Greenberg, B. J Clin Psych. 1998, 59, 4-12 6. Bengel, D.;Murphy, D.L.; Andrews, A.M.; Wichems, C.H.; Feltner, D.; Heils, A.; Mossner, R.; Westpal, H.; Lesch, K.P. Molecular Pharmacolpogy. 1998, 53, 649-655 Homozygote SERT Knockout Mice Figure 4. DA Uptake by Chronoamperometry Representative DA electrochemical signal from SERT knockout mice striatal synaptosomes after the addition of 1 M DA. Only oxidation signals are illustrated.