Download
1 / 53
530 likes | 814 Vues
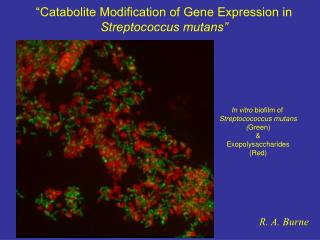
“Catabolite Modification of Gene Expression in Streptococcus mutans”. In vitro biofilm of Streptocococcus mutans ( Green) & Exopolysaccharides (Red). R. A. Burne. Streptococcus mutans…. Primary causative agent of dental caries Classic Biofilm Disease

E N D
“Catabolite Modification of Gene Expression in Streptococcus mutans” In vitro biofilm of Streptocococcus mutans (Green) & Exopolysaccharides (Red) R. A. Burne
Streptococcus mutans… Primary causative agent of dental caries Classic Biofilm Disease Significant contributor to infectious endocarditis “Obligate” biofilm lifestyle No free-living existence
Mechanisms of Biofilm Formation by S. mutans Sucrose-Independent Multiple Cell-surface Associated Adhesins P1 (SpaP) is primary adhesin Binding to salivary receptors (gp340, statherin, mucin) Sucrose-Dependent Facilitated by production of glucan homopolymers Products of the Glucosyltransferases (GTFs) a1,3- & a1,6-linked glucans Multiple glucan binding proteins (3-4 + GTFs) Particularly important on smooth surfaces
Exopolysaccharide Metabolism Overview
Fructan exopolysaccharides increase the depth and duration of biofilm acidification 1 2 4 3
Fructan exopolysaccharides increase the depth and duration of biofilm acidification 1 2 4 3
Fructan exopolysaccharides increase the depth and duration of biofilm acidification 1 2 4 3
Fructan exopolysaccharides increase the depth and duration of biofilm acidification 1 2 4 3
Characteristics of FruA 1. Secreted 2. Levans > Inulins > Sucrose > Raffinose (b2,6) (b2,1) (b2,1) (b2,1) HMW HMW di-Sac gal-sucr 3. Exo-hydrolase Releases fructose from b-fructosidases 4. FruA-deficient strains have diminished virulence in a rat model
Substrate pH 5.5 + C N fruA pH 7.0 - CCR Complex control of FruA + Growth Rate, pH, O2, other
Primer Extension TIS ATG DS Pfru 166 bp CRE-W CRE-S RBS “Extended -10 Promoter Terminator + RAT The fruA Promoter Region
Summary - cis-acting elements Induction by subtrate is mediated through a dyadic sequence located at -59 relative to the TIS. Repression is mediated by two catabolite response elements with high conservation with CcpA binding sites. The leader mRNA plays a relatively minor role in fruA expression, perhaps by affecting mRNA stability. ATG TIS DS 166 bp Pfru CRE-W CRE-S RBS “Extended -10 Promoter Terminator + RAT
UA159 Genome Ajdic et al, PNAS 22:14434-14439 How is fruA induced by its substrates?
Signal peptide Transmembrane domain A “four-component system” encoded in the S. mutans genome
The TCS and sugar binding proteins are required for fruA expression WT TCS, SBPs Growth on Inulin (2,1-linked fructan)
levT levR levS levQ The phosphoenolpyruvate-dependent sugar: phosphotransferase system (PTS) levG levF levE levD
CAT Activity fruA expression is enhanced in the levD mutant
Conclusion I An unusual “four-component” signal transduction system is required for activation of fruA The most potent activating signal appears to be fructose, but for reasons not fully understood, mannose is as efficient The LevDEFG fructose/mannose PTS permease is a negative regulator of fruA, extending the role of the PTS in governance of fructan catabolism Fine-structure mutagenesis is beginning to identify key sequence:function relationships in the LevQRST system
Carbohydrate Catabolite Repression in Streptococcus mutans Deviation from the paradigm?
Catabolite Repression in Gm (+) Bacteria FROM: Deutscher J, Herro R, Bourand A, Mijakovic I, Poncet S. Biochim Biophys Acta. 2005 Dec 30;1754(1-2):118-25.
Wen & Burne Loss of CcpA did not alleviate CCR in cells growing on inulin and glucose
Dissecting the induction pathway allowed for exploration of CCR by pulsing with LevQRST substrates
Therefore, CcpA does exert CCR over the fruA operon, but redundancies exist…probably allowing for thresholding of repression
Interaction of CcpA with the fruA promoter is direct and occurs through the CRE
88 genes differentially expressed at p < 0.005 Transcription Profiling Wild-type versus ccpA-
Conclusions - CcpA & CCR CcpA does in fact exert control of CCR in S. mutans CcpA plays a role in global control of gene expression, functioning both as a repressor and activator of transcription in response to CHO CcpA governs the expression of multiple virulence attributes and loss of CcpA affects many virulence-related phenotypes S. mutans possesses redundant systems for CCR to fine-tune gene expression in response to CHO sources and availability
A A B B PEP IIman PO4 C EI PO4 HPr IIfru A IIlev Pyruvate Progress on CCR - Role of the PTS
ManL (IIman) mediates repression in response to mannose and glucose, not fructose
FruI (IIfru) mediates repression in response to fructose and glucose, not mannose
LevD (IIlev) mediates repression in response to mannose and fructose
Loss of CcpA enhances repression by mannose and fructose Because…
F/M LevT LevQ LevT LevQ F/M G LevS LevS P ATP ADP M CcpA P LevR F FruI LevR G/M X EIIMan fruA LacR, CelR etc. P P levDE fruA levDE CcpA F F/M F EIILev F F
S. mutans UA159 eluate from a Ni-NTA column 4 7 kDa 200 116.3 97.4 66.3 55.4 36.5 31 21.5 14.4 UA 159 MB-08-19
Ni-NTA eluate from S. mutans expressing histidine-tagged ManL 4.0 5.4 7.0 kDa 200 116.3 97.4 66.3 55.4 36.5 31 21.5 14.4 GLUCOSE
Ni-NTA eluate from S. mutans expressing histidine-tagged ManL 4 7 200 116.3 97.4 66.3 55.4 36.5 31 21.5 14.4 Fructose
F/M LevT LevQ LevT LevQ F/M G LevS LevS P ATP ADP M CcpA P LevR F FruI LevR G/M X EIIMan fruA LacR, CelR etc. P P levDE fruA levDE CcpA F F/M F EIILev F F
Structure:function analysis of the levQRST genes Progress to date GPS™-LS Linker Scanning System
K434x1 I430x3 A113 P157 G210 I426x3 V419 P152 K391x1 E66 N338x1 L409x2 F65x1 T313x1 G96x1 Q350x1 S314x2 G351x1 F134 K391x1 I430x4 K389x1 L414x1 I426x2 E229 K326x1 I402x1 D194x4 N360 F346 V74x2 L250x1 A307 H69x1 No growth on TVI K395x1 L181x1 Y254x4 F261x1 L414x5 G197x1 W.T. level Growth on TVI K412x1 F331x1 F261x1 T411x1 0 100 200 300 400 435 Increased Growth on TVI GPS-linker scanning of LevQ