Download
1 / 45
450 likes | 469 Vues
Explore the reorganization and function of the hippocampus in memory formation across different mammalian species, including humans, rats, monkeys, and birds. Learn about the role of the hippocampus in spatial and episodic memory, and the potential for content-addressable memory. Discover how associative networks and plasticity levels contribute to memory storage and retrieval in the hippocampus.

E N D
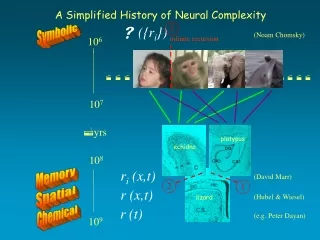
r (t) Chemical (e.g. Peter Dayan) 106 r (x,t) Spatial (Hubel & Wiesel) ri (x,t) Memory (David Marr) 107 2 3 1 yrs platypus echidna DG 108 CA3 CA1 lizard ({ri}) Symbolic (Noam Chomsky) 109 A Simplified History of Neural Complexity infinite recursion mammalian species
Let us concentrate on one stepon our way out of the primordial neural tube • first, a process of evagination
and then… some slight change in our chips:
and then… some slight change in our chips:
and then… some slight change in our chips: • the medial wall reorganized • into the hippocampus
the reorganization includes a spatial migration... ...it does not lead to a new type of cortex...
but the reorganization of the hippocampus... ….occurs, fundamentally, through the detachment and granulation of the dentate gyrus (diagrams taken from the book by P. Gloor)
it is, at its core, a granulation! …self-similar across species
watch it frozen in its development, in the opossum
David Marr, over 30 years ago, suggested to start from the function In humans, the hippocampus had long been implicated in the formation of episodic and autobiographical memories (here, data by Graham & Hodges)
Over the last few years, imaging evidence has corroborated traditional neuropsychological evidence (here, fMRI study of verbal encoding into episodic memory by Fernandez et al)
In rats, the evidence from neurophysiological recordings indicates a primary role in spatial memory (here, data from simultaneous recordings by Matt Wilson & Bruce McNaughton)
(although a minority view has emphasized a more active role in spatial computation; here, data by Neil Burgess & John O’Keefe)
In monkeys, Edmund Rolls et al have found spatial view cells, suggestive of a hippocampal role intermediate between the human and the rat description
The extensive extrinsic connections of the hippocampus with the neocortex are consistent with a role in the formation of memories
David Marr’s perspective was the same lateradopted by most of his followers... (diagram by Jaap Murre, 1996)
So, let us follow the same functional hypothesis... …but let us try to be quantitative
What sort of device can…. • generate, on line, appropriate (compressed) representations of each “snapshot” • store these representations on line, in a single “shot” • hold multiple representations simultaneously • retrieve each representation from partial cues • send back the retrieved information in a robust format ?
generate, on line, appropriate (compressed) representations of each “snapshot” • store these representations on line, in a single “shot” • hold multiple representations simultaneously to retrieve each representation from partial cues • send back the retrieved information in a robust format • requires a content addressable memory CAM Icue=log2p << Iitem Niunit
The analysis of large- scale recordings (here, by Skaggs & McNaughton) shows that the information content of hippocampal representations grows linearly with population size, before saturating at the ceiling set by the experiment. Francesco Battaglia has quantified the full Iitem for place cells, using an analytical model.
generate, on line, appropriate (compressed) representations of each “snapshot” to store representations on line, in a single “shot”, • hold multiple representations simultaneously • retrieve each representation from partial cues • send back the retrieved information in a robust format (with neuronally plausible mechanisms) • is something Hebbian associative networks can do ... p ~ 0.2 C / [a log(1/a)] (Francesco has shown that with maps it is just the same)
If LTP modifies the same synapses affected by learning... one may predict the effect of lower C values higher dp/dt
generate, on line, appropriate (compressed) representations of each “snapshot” • to store representations on line, in a single “shot” multiple representations • retrieve each representation from partial cues • send back the retrieved information in a robust format • are held most efficiently in a free autoassociator, which minimizes the components required for a given information content
generate, on line, appropriate (compressed) representations of each “snapshot” • store these representations on line, in a single “shot” • hold multiple representations simultaneously • retrieve each representation from partial cues the read-out of the retrieved information • is greatly facilitated by expansion recoding with additional associative “polishing”
Analytical models predict an optimal plasticity level for CA3->CA1 (Schaffer) collaterals, but are not yet constrained enough to predict the observed memory activation differences CA3 CA1
…but, why do we need CA1, then? the answer may lie in the predictive ability that several models assign to the hippocampus (as well as to any associative network with time-asymmetric plasticity) although CA3 may predict future “contexts” as well as CA1, this may conflict with devoting its recurrent collaterals to retrieve the current “context”. We shall get back to this issue tomorrow…
generating information-rich compressed representations • store these representations on line, in a single “shot” • hold multiple representations simultaneously • retrieve each representation from partial cues • send back the retrieved information in a robust format • requires a dedicated preprocessor that sparsifies and decorrelates input activity
PP inputs (from EC) modify during storage and relay the cue at retrieval Why the separation of a dentate gyrus? MF inputs (from DG) force an informative representation at storage and are irrelevant for retrieval
The crucial prediction is consistent with recordings from normal rats
(Tucson data by Jim Knierim) but it is difficult to test it in dentate lesioned rats
Acquisition Index: Errors (T1-5D1) - (T6-10D1) Retrieval Index: Errors (T6-10D1) - (T1-5D2) Double dissociation





















