Speciation Dynamics of an Agent-based Evolution Model in Phenotype Space
390 likes | 516 Vues
This study explores speciation dynamics through an agent-based evolution model that operates within phenotype space, where phenotypes reflect genetic traits. It examines clustering and phase transitions on neutral landscapes, mechanisms of mutation, and their implications on species diversity. The model addresses the principles of sympatric speciation in various organisms, including bacteria and fungi, and incorporates factors such as assortative mating and fitness landscapes. Key findings suggest critical roles of mutability and reproductive strategies in shaping evolutionary outcomes.

Speciation Dynamics of an Agent-based Evolution Model in Phenotype Space
E N D
Presentation Transcript
Speciation Dynamics of an Agent-based Evolution Model in Phenotype Space Adam D. Scott Center for Neurodynamics Department of Physics & Astronomy University of Missouri – St. Louis Oral Comprehensive Exam 5*31*12
Proposed Chapters • Chapter 1: Clustering and phase transitions on a neutral landscape (completed) • Chapter 2: Simple mean-field approximation to predict universality class & criticality for different competition radii • Chapter 3: Scaling behavior with lineage and clustering dynamics
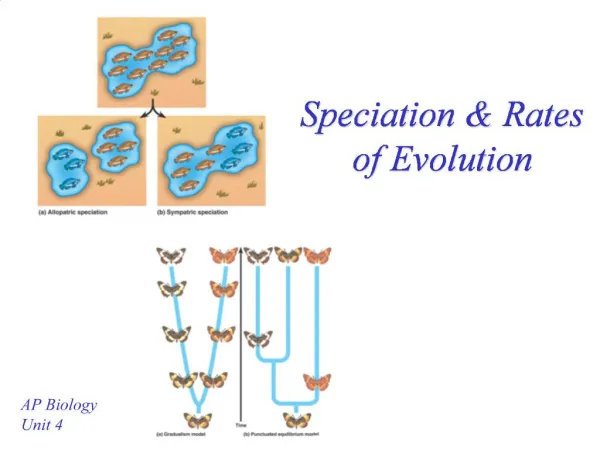
Basis Biological • Modeling • Phenotype space with sympatric speciation • Phenotype = traits arising from genetics • Sympatric = “same land” / geography not a factor • Possibility vs. prevalence • Role of mutation parameters as drivers of speciation • Evolution = f(evolvability) • Applicability Physics & Mathematics • Branching & Coalescing Random Walk • Super-Brownian • Reaction-diffusion process • Mean-field & Universality • Directed &/or Isotropic Percolation
Broader Context/ Applications • Bacteria • Example: microbes in hot springs in Kamchatka, Russia • Yeast and other fungi • Reproduce sexually and/or asexually • Nearest neighbors in phenotype space can lead naturally to assortative mating • Partner selection and/or compatibility most likely • MANY experiments involve yeast
Model: Overview • Agent-based, branching & coalescing random walkers • “Brownian bugs” (Young et al 2009) • Continuous, two-dimensional, non-periodic phenotype space • traits, such as eye color vs. height • Reproduction: Asexual fission (bacterial), assortative mating, or random mating • Discrete fitness landscape • Fitness = # of offspring • Natural selection or neutraldrift • Death: coalescence, random, & boundary
Model: “Space” • Phenotype space (morphospace) • Planar: two independent, arbitrary, and continuous phenotypes • Non-periodic boundary conditions • Associated fitness landscape
Model: Fitness Natural Selection Neutral Theory Hubbell Ecological drift Kimura Genetic drift Equal (neutral) fitness for all phenotypes No deterministic selection Random drift Random selection Fitness = 2 • Darwin • Varying fitness landscape over phenotype space • Selection of most fit organsims • Applicable to all life • Fitness = 1-4 • (Dees & Bahar 2010)
Model: Mutation Parameter • Mutation parameter -> mutability • Ability to mutate about parent(s) • Maximum mutation • All organisms have the same mutability • Offspring uniformly generated Example of assortative mating assuming monogamous parents
Model: Reproduction Schemes • Assortative Mating • Nearest neighbor is mate • Asexual Fission • Offspring generation area is 2µ*2µ with parent at center • Random Mating • Randomly assigned mates
Model: Death • Coalescence • Competition • Offspring generated too close to each other (coalescence radius) • Random • Random proportion of population (up to 70%) • “Lottery” • Boundary • Offspring “cliff-jumping”
Model: Clusters • Clusters seeded by nearest neighbor & second nearest neighbor of a reference organism • A closed set of cluster seed relationships make a cluster = species • Speciation • Sympatric Cluster seed example: The white organism has nearest neighbor, yellow (solid white line). White’s 2nd nearest neighbor is blue (hashed white line). Therefore, white’s cluster seed includes: white, yellow, and blue.
Generations 00.40 1 50 1000 2000 00.44 µ 00.50 01.20
Chapter 1: Neutral Clustering & Phase Transitions • Non-equilibrium phase transition behavior observed for assortative mating and asexual fission, not for random mating • Surviving state clustering observed to change behavior above criticality
Assortative Mating • Potential phase transition • Extinction to Survival • Non-equilibrium • Extinction = absorbing • Critical range of mutability • Large fluctuations • Power-law species abundances • Peak in clusters Quality (Values averaged over surviving generations, then averaged over 5 runs)
Asexual Fission • Slightly smaller critical mutability • Same phase transition indicators • Same peak in clusters • Similar results for rugged landscape with Assortative Mating
Control case: Random mating Generations 1 50 1000 2000 02.00 µ 07.00 12.00
Random Mating • Population peak driven by mutability & landscape size comparison • No speciation • Almost always one giant component • Local birth not guaranteed!
Conclusions • Mutability -> control parameter • Population as order parameter • Continuous phase transition • extinction = absorbing state • Directed percolation universality class? • Speciation requirements • Local birth/ global death (Young, et al.) • Only phenotype space (compare de Aguiar, et al.) • For both assortative mating and asexual fission
Chapter 1: Progress • Manuscript submitted to the Journal of Theoretical Biology on April 16 • Under review as of May 2 • No update since
Chapter 2 • Goal: to have a tool which predicts critical mutability and critical exponents for a given coalescence radius = Mean-field equation • Directed percolation (DP) & Isotropic percolation (IP) • Neutral landscape with fitness = 2 for all phenotypes • May extend to arbitrary fitness if possible • Asexual reproduction • Will attempt extension to assortative mating
Temporal & Spatial Percolation • Temporal Survival • Time to extinction becomes computationally infinite • DP • Spatial “Space filling” • Largest clusters span phenospace • IP
1+1 Directed Percolation • Reaction-diffusion process of particles • Production: A2A • Coalescence: 2AA • Death: A0 • Offspring only coalesce from neighboring parent particles N N+1 Death (A→ᴓ) Production(A→2A) Coalescence (2A →A)
Chapter 2: Self-coalescence • Not explicitly considered in basic 1+1 DP lattice model • Mimics diffusion process • May act as a correction to fitness, giving effective birth rate • “Sibling rivalry” • Probability for where the first offspring lands in the spawn region • Probability that the second offspring lands within a circle of a given radius whose center is offspring one and its area is also in the spawn region 2 1
Chapter 2: Neighbor Coalescence • Offspring from neighboring parents coalesce Coalescence (2A →A) 2 1 1 2
Assuming Directed Percolation • Simple mean-field equation (essentially logistic) • Density as order parameter • τ is the new control parameter • should depend on mutability and coalescence radius • is effective production rate (fitness & self-coalescence) • is effective death rate (random death) • g is a coupling term • g = , the effective coalescence rate (”neighbor rivalry”)
Chapter 2: Neutral Bacterial Mean-field • Birth: • Coalescence: • Random death: • Effective production rate = • Effective death rate = • Effective coalescence rate = ? • Possibly a coupled dynamical equation for nearest neighbor spacing • & • Without nc, current prediction for critical mutability (~0.30) is <10% from simulation (~0.33)
Chapter 2: Neighbor Coalescence • Increased rate with larger mutability & coalescence radius • Varies amount of overlapping space for coalescence • Should depend explicitly on nearest neighbor distances • May be determined using a nearest neighbor index or density correlation function • Possibility of a second dynamical equation of nearest neighbor measure coupled with density?
Chapter 2: Progress • Have analytical solution for sibling rivalry • Have method in place to estimate neighbor rivalry • Waiting for new data for estimation • Need to finish simple mean-field equation • Need data to compare mean-field prediction of criticality for different coalescent radii • Determine critical exponents • Density, correlation length, correlation time
Chapter 3: Scaling • Can organism behavior predict lineage behavior? • Center of “mass” center of lineage (CL) • Random walk • Path length of descendent organisms & CL • Branching & (coalescing) behavior • Can organism behavior predict cluster behavior? • Center of species (centroids) • Clustering clusters • Branching & coalescing behavior • May determine scaling functions & exponents • Population # of Clusters? • Fractal-like organization at criticality? • Lineage branching becomes fractal? • Renormalization: organisms clusters
Chapter 3: Cluster level reaction-diffusion • Clusters can produce n>1 offspring clusters • AnA (production) • Clusters go extinct • A0 (death) • m>1 or more clusters mix • mAA (coalescence)
Chapter 3: Predictions • Difference of clustering mechanism by reproduction • Assortative mating: organisms attracted (sink driven) • Greater lineage convergence (coalescence) • Bacterial: clusters from blooming (source driven) • Greater lineage branching (production) • Greater mutability produces greater mixing of clusters & lineages • Potential problem: far fewer clusters for renormalization
Chapter 3: Progress • Measures developed for cluster & lineage behavior • Extracted lineage and cluster measures from previous data • Need to develop concrete method for comparing the BCRW behavior between reproduction types • ?
Related Sources • Dees, N.D., Bahar, S. Noise-optimized speciation in an evolutionary model. PLoS ONE5(8): e11952, 2010. • de Aguiar, M.A.M., Baranger, M., Baptestini, E.M., Kaufman, L., Bar-Yam, Y. Global patterns of speciation and diversity. Nature460: 384-387, 2009. • Young, W.R., Roberts, A.J., Stuhne, G. Reproductive pair correlations and the clustering of organisms. Nature412: 328-331, 2001. • HinsbyCadillo-Quiroz, Xavier Didelot, Nicole Held, Aaron Darling, Alfa Herrera, Michael Reno, David Krause and Rachel J. Whitaker. Sympatric Speciation with Gene Flow in Sulfolobusislandicus.PLoS Biology, 2012. • Perkins, E. Super-Brownian Motion and Critical Spatial Stochastic Systems. http://www.math.ubc.ca/~perkins/superbrownianmotionandcriticalspatialsystems.pdf. • Solé, Ricard V. Phase Transitions. Princeton University Press, 2011. • Yeomans, J. M. Statistical Mechanics of Phase Transitions. Oxford Science Publications, 1992. • Henkel, M., Hinrichsen, H., Lübeck, S. Non-Equilibrium Phase Transitions: Absorbing Phase Transitions. Springer, 2009.
µ = 0.38 µ = 0.40 slope ~ -3.4 • Power law distribution of cluster sizes • Scale-free • Large fluctuations near critical point (Solé 2011) • Characteristic of continuous phase transition • Near criticality parabolic distributions change gradually • Mu < critical concave down • Mu > critical concave up µ = 0.42
Clark & Evans Nearest Neighbor Test Asexual Fission Clustered <= 0.46 (peak) Dispersed >= 0.54 Better than 1% significance Assortative Mating • Clustered <= 0.38 (peak) • Dispersed >= 0.44 • Better than 1% significance