Competition
Competition. Peter B. McEvoy Insect Ecology Ent 420/520. Prevalence of current competition in natural communities as judged by recent reviews. Schoener (1983) Connell (1983) Strong, Lawton, Southwood (1984) Lawton and Hassell (1984) Denno, McClure, Ott (1995). Review by Denno et al. .


Competition
E N D
Presentation Transcript
Competition Peter B. McEvoy Insect Ecology Ent 420/520
Prevalence of current competition in natural communities as judged by recent reviews • Schoener (1983) • Connell (1983) • Strong, Lawton, Southwood (1984) • Lawton and Hassell (1984) • Denno, McClure, Ott (1995)
Review by Denno et al. • History of Competition studies • Failure to find evidence of competition probably lies with the methods • Observational studies of pattern (e.g. niche and habitat overlap) were inappropriately used to infer process. Is high overlap indicative of presence or absence of competition? • Experimental studies of competition in phytophagous insects were few and far between (e.g. in reviews by Connell and Schoener) • Recent application of more powerful methods
Denno, R. F., M. S. McClure, and J. R. Ott. 1995. Interspecific interactions in phytophagous insects: Competition reexamined and resurrected. Ann. Rev. Entomol. 40:297-331. • Scope. 193 pair-wise species interactions (all major feeding guilds except pollinators and detritivores): occurrence, frequency, symmetry, consequences, and mechanisms of competition. • Result. Interspecific competition in 76% of interactions, often asymmetric, and frequent in most guilds (sap feeders, wood and stem borers, seed and fruit feeders) except free-living mandibulate folivores.
Review Continued • Phytophages more likely to compete if they were closely related, introduced, sessile, aggregative, fed on discrete resources, and fed on forbs or grasses • Interference competition was most frequent between mandibulate herbivores living in concealed niches • Host plants mediated competitive interactions more frequently than natural enemies, physical factors, and intraspecific competition
Narrative and vote-counting reviews can be misleading… • Move from • Narrative or qualitative reviews • Vote-counting reviews that assess the frequency of reported outcomes • Such comparisons can be misleading because they fail to consider the magnitude of treatment effects and ignore the importance of sample size in determining the statistical power of individuals studies • Meta-analysis offers a remedy
Paradigms Lost • Top Down Effects • Herbivores rarely compete • Herbivores rarely influence plant population and community dynamics • Bottom Up Effects • Much of the green is inedible or unsuitable • Some downright toxic C H P
Effects of Carnivore RemovalMeta-analysis by Schmitz et al. 2000 • Direct positive effect of carnivore removal on herbivores • Indirect negative effect of carnivore removal on plants • Plant biomass • Feeding Damage • Reproductive output (seeds) C H P
Effects of Herbivore RemovalMeta-analysis by Bigger and Marvier 1998 A meta-analysis of 246 experimental contrasts between plants protected from herbivores versus plants exposed to herbivores Herbivores often impose important effects on plant biomass -- equal in magnitude to those imposed by plant competition. Effects of invertebrate herbivores are significantly stronger than those of vertebrates, in contrast to widely held views. Thus future studies should investigate how herbivory and plant competition interact to determine the distribution, abundance, and diversity of plants. H P
Persistent Questions • Are herbivores limited by natural enemies? …to such a degree that they rarely compete or reduce plant biomass? • Are herbivores limited by food resources? Quantity or Quality? What are the elements of Host Quality? • Are plants limited by herbivores?
Rules of the Game are ChangingHumans have Transformed Constraints on Plant Growth C • Resource limitation • Recruitment limitation • Predators and pathogens • Disturbance • Conditions (temperature/climate) • Temporal variation H P R
Paradigms GainedExperiments on CompetitionReview by Denno, McClure, Ott 1995 • Is competition mediated by • Carnivores • Diffuse interactions among many herbivores • Changes in plant quality C H P
Concept Alert! • Competition and Coexistence • Character displacement • Resource partitioning • Exploitation vs Interference • Coevolution of competitors – interaction a prerequisite • Null hypothesis: unsaturated, non-interactive communities
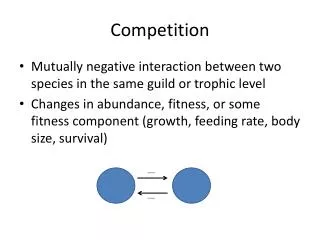
Concept Alert! Mechanisms of Competition • Exploitative competition occurs when individuals, by using resources, deprive others of the benefits to be gained from those resources • Interference Competition results when individual harm one another directly through fighting or killing, or indirectly by aggressively maintaining a territory or producing chemicals that deter other individuals
Asymmetric Competition • For equal competitors, strong intraspecific competition may mitigate interspecific competition • For asymmetric competitors, strong intraspecific effects in the superior competitor may not preclude a strong interspecific effects on the inferior competitor
Key Questions • Conditions for coexistence. What conditions lead to coexistence vs. competitive exclusion? • Frequency and intensity. How common is interspecific competition in nature, what kinds of species are likely to compete? • Niche differentiation. What is extent of niche differentiation among coexisting competitors? Is competition a cause of morphological and ecological differentiation among species?
Predictions That Follow From Conventional Theory • Potential competitors should exhibit niche differentiation • Niche differentiation often manifest as morphological differentiation • With any community, species with little or no niche differentiation are unlikely to coexist (limiting similarity), likely to differ in habitat (habitat distribution)
Niche Separation Along a Resource Axis Narrow niches with little overlap (d > w) Broad niches with greateroverlap (d < w)
Conditions for Coexistencerelated to carrying capacities K1 and K2 and nich overlap (d/w) Decreasing d/w (increasing overlap) reduces likelihood of stable coexistence except for competitors with similar carrying capacities
Morphological Differentiation Among Harvester Ants As species diversity of ants increases, the variability in mandible length for Veromessor pergandei decreases
Character Displacement in Harvester Ants Observational studies Mandible sizes of this species vary from site to site such that they always differ from those of local competitors (shown by arrows)
History of Competitive Displacement in Aphytis Parasitoids • Aphytis chrysomphali • A. lingnanensis • A. melinus Displaced by Displaced by
Mechanism of Competitive Displacement in Aphytis Minimum host size required for female progeny is larger for inferior competitorA. lingnanesis (right arrow) than for superior competitor A. melinus (left arrow) for red scale distributions on wood, leaves, fruits
Females of 4 Species of Parasitoids Attacking Cocoon Stage of Swaine Jack Pine Sawfly Note differences in wing area and ovipositor length
Ratios in Ovipositor Lengths in Parasitoids Attacking Sawfly Pupae: Too Close for Coexistence?
Methodological difficulties in proving or disproving Competitive Exclusion principle • When two species compete and coexist, we can suppose that they do so as a result of some niche differentiation. • But this supposition remains unconfirmed until such differentiation is observed and shown to ameliorate the effects of interspecific competition. • It is often difficult to establish that there is such differentiation, and it is impossible to establish the absence of it. We may simply have looked in the wrong place or in the wrong way.
Modern theory moving away from assumption of community at equilibrium • Disturbance, colonization, successional development • Unpredictable gaps: the poorer competitor is a better colonizer (Zwölfer 1973) • Unpredictable gaps: the preemption of space • Fluctuating environments: independence in species fluctuations • Ephemeral patches with variable life-spans: the poorer competitor reproduces more rapidly • Aggregated distributions - e.g. carrion flies (Ives 1988, Ives 1991) • Apparent competition: coexistence mediated by a shared natural enemy • No effect on coexistence. Predation modifies the interaction between competitors without necessarily making coexistence more likely • Promotes coexistence. Predation more likely to enhance diversity if predator prefers to feed on the superior competitor
Effects of Species Interactions on Biological InvasionsApparent Competition in Leafhoppers(Settle and Wilson 1990) Parasitoid Grape Leaf Hopper Variegated leaf hopper Resident Herbivore Invading Herbivore VLH GLH Host Plant Grape
Relative Strength of Intraspecific and Interspecific Competition • Density-dependent per capita reproduction • Equivalence of intraspecific and interspecific competitors indicated by equal slopes
“Apparent Competition” or Negative Effect of One Species on Another Acting Via a Shared Parasitoid • Percent GLH parasitism • Ratio of parasitoids to GLH Increase with VLH relative density
Non-linear, frequency-dependent parasitism Because risk of parasitism is higher in GLH than VLH, VLH contributes a small fraction of parasitoids even when VLH is an intermediate fraction (i.e. up to 50%) of the total hopper population but contribution accelerates thereafter
Populus angustifolia(Narrowleaf Cottonwood) • http://www.public.iastate.edu/~bot356/species/species/p_tSpecie/PopuAngu.html
Intraspecific Competition in Pemphigus Aphids studied by Whitham • Narrow “window” for colonization in spring. As much as 83% of over-wintering Pemphigus betae population migrates in 3-day span to immature leaves of Populus angustifolia were they gall and become encased in plant tissue
Habitat selection by Pemphigus aphids in response to resource limitation and competition Successfully developed gall of Pemphigus betae on leaves of Populus angustifolia Aborted gall
Size of mature leaf is critical to ultimate fitness of individual colonizing mother (A) Percent aborted declines… (B) Aphids per gall increases… (C ) Stem mother weight increases.. …with increasing leaf size
Percent Stem mothers seek out and colonize large leaves Distribution of Leaves Available on Tree Distribution of Leaves used by stem mothers
Territorial AphidsTypical fighting posture of Pemphigus betae females. The larger female usually wins the better territory, which is about 3 mm long at the base of the leaf Whitham 1979
Intraspecific competition in females of the aphid Pemphigus betaeWhen dominant female removed, displaced female moved down to occupy the better position Whitham 1980
When more than one aphid colonizes a leaf, the advantage goes to the aphid lower on the leaf • Effect of gall position and numbers of aphids on females fitness • Leaves drawn to scale and effects of position given in boxes • Trends in leaf size, aphids per gall, abortion rate, stem mother weight
Model of Habitat Selection • The Fretwell and Lucas model of habitat choice assumes that fitness is negatively correlated with density of competitors. • As best habitat is colonized and densities become high, poorer but sparsely inhabited habitats are likely to be used • Fitness similar across habitats. Prime habitat with more occupants becomes similar to poorer habitat with fewer occupants At d2 At d3
Fretwell Lucas model applied to Pemphigus betae aphids • Shows habitats (leaves) of varying quality (leaf size) and differences in population density per habitat (galls per leaf) • Lines show expected fitness (aphids per gall) among leaf sizes and gall densities. • Solid line indicates how average fitness for 1,2,3 stem mothers per leaf is even across densities, consistent with model predictions
Points made Not all leaves on tree are available or suitable When available resources accurately defined, found to be limiting at least within tree (we don’t yet know tree to tree picture) Points requiring further study Host variability as defense against herbivores Intertwining of suitable factors: nutrition, density of conspecifics, leaf abscission, enemies Conclusions from Pemphigus study
Insects on Seaside Daisy • Erigeron glaucus Seaside daisy Platyptilia emissalisPTEROPHORIDAE is a congener of species studied by Karban
Herbivore Example: Spittlebugs, Plume Moths, and thrips on Seaside Daisy (Karban) • Combines experimental and observational study of interspecific competition between folivorous insects on seaside daisy Erigeron glaucus • The study illustrates asymmetric competition between unrelated insects belonging to different feeding guilds
Natural History • The Calendula plume moth Platyptilia williamsii (Lepidoptera: Pterophoridae) is a multivoltine, shoot-feeder. • The meadow spittlebug Philaenus spumarius (Homoptera: Cercopidae) is a cosmopolitan, univoltine, sap-tapper with a very broad host plant range.
Experimental Design • Treatments manipulated the density of each insect: 2 levels of moths (presence, absence) x 2 levels of spittlebugs. Presence was represented by a single fixed density for each insect (1 moth, 5 spittlebugs). Would have been better to have 3 levels of each factor. Why? • Responses were measured as persistence of each insect and approximate population growth rates.
Resource Renewal Rates and Competition Mechanisms • Leaf-axils are a limiting resource for spittlebugs (McEvoy 1986) and mechanical barriers to feeding confer resistance (Hoffman and McEvoy 1985, Hoffman and McEvoy 1986). • Karban measured resource renewal rates (production of new leaves), simulated effects of plume moth by binding leaves together, and found similar results with simulated and observed damage.
Multiple Scales of Observation • Important to look across scales, see how description of competition changes with scales, and to link processess occuring on different scales • Karban investigated multiple scales • temporal scales (Within Between Years) • spatial scales (Plant PatchRegional Patterns of Association) • organizational scales (Two focal species in this paper Other species including a thrips, a leafhopper, and an aphid are treated in other papers).
Results Insects on seaside daisy • Asymmetric interactions. Persistence times and net reproductive rates of spittlebugs are reduced by addition of plume moth; but there is no detectable effect of spittlebugs on plume moths • Individualistic behavior. Each species affected strongly by a different biotic factor • Spittlebugs by interspecific competition • Plume moths by vertebrate predation • Thrips by clonal variation