Gene duplication, exon shuffling and concerted evolution
540 likes | 2.13k Vues
Gene duplication, exon shuffling and concerted evolution. Xuhua Xia xxia@uottawa.ca http://dambe.bio.uottawa.ca. Evolution of new functions. New function through lateral gene transfer Transformation Transduction Conjugation New function through sequence duplication: Gene duplication

Gene duplication, exon shuffling and concerted evolution
E N D
Presentation Transcript
Gene duplication, exon shuffling and concerted evolution Xuhua Xia xxia@uottawa.ca http://dambe.bio.uottawa.ca
Evolution of new functions • New function through lateral gene transfer • Transformation • Transduction • Conjugation • New function through sequence duplication: • Gene duplication • Internal domain duplication • Exon/domain shuffling • New function through alternative processing • Alternative splicing • RNA editing • Overlap genes (alternative transcription) • Different mixtures of gene products How do complex organisms acquire extra genes (for new functions)? … and extra forms of regulation? Slide 2
Horizontal Gene Transfer (P. 359-366) - lateral transfer of genetic information from one genome to another (eg. between two species) Mechanisms: • Transformation • - via free DNA (vector not essential) 2. Transduction - via bacteriophage or virus 3. Conjugation in bacteria - via conjugative plasmid Estimated that ~ 10-18% of E.coli genome due to LGT eg. lactose operon (milk sugar lactose used as carbon source in mammalian colon) (Starlinger 1977; Riley and Anilionis 1980; Buvinger et al. 1984; Ochman, Lawrence, and Groisman 2000; Lawrence 2001, but see Stoebel, D. M., 2005. Lack of Evidence for Horizontal Transfer of the lac Operon into Escherichia coli. Mol Biol Evol 22, 683-690.). Slide 3
Inferred HGT of lactose operon • Inference: lactose operon is acquired through HGT • Prediction: genes in lactose operon should not reflect true phylogenetic relationship. Stoebel, D. M. Mol Biol Evol 2005 22:683-690; doi:10.1093/molbev/msi056 Slide 4
Trees from house keeping genes Stoebel, D. M. Mol Biol Evol 2005 22:683-690; doi:10.1093/molbev/msi056 Slide 5
Trees based on lacZ and lacZY data Stoebel, D. M. Mol Biol Evol 2005 22:683-690; doi:10.1093/molbev/msi056 Slide 6
Acid-resistant mechanisms Stomach fluid:Extrinsic source ureAB-I-EFGH Nickel- inserting proteins Urea Urease (A+B) apoenzyme Arginine+arginase:Intrinsic source Ammonia Slide 7
Evolution of new functions • New function through lateral gene transfer • Transformation • Transduction • Conjugation • New function through sequence duplication: • Gene/genome duplication • Internal domain duplication • Exon/domain shuffling • New function without sequence duplication: • Alternative splicing • RNA editing • Overlap genes (alternative transcription) • Different mixtures of gene products How do complex organisms acquire extra genes (for new functions)? … and extra forms of regulation? Slide 8
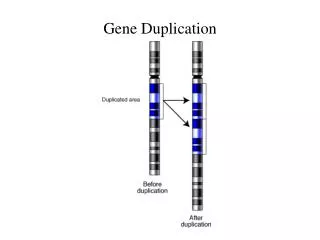
New genes through duplication Hartwell Fig. 21.11 Slide 9
1. Gene duplication - one copy can perform original function and second one may evolve new function (or degenerate into pseudogene) Tandem arrays chr 1 Dispersed copies chr 5 Multi-gene families – sets of genes derived by duplication of ancestral gene Pseudogene – non-functional member of gene family Slide 10
Homologues and paralogues Homologous genes - share common evolutionary origin Orthologous genes • descendants of an ancestral gene that was present • in the last common ancestor of two or more species Paralogous genes - arose by gene duplication within a lineage Species 2 Species 1 Species 3 ancestor Slide 11
Gene duplication and gene phylony A1H A1C A1 A1H A2H A1 A2 A1 A1 A1 A2 A1M A1R A1 A1 A1C A2C A1 A1 A2 A1 A1 A A A2 A2 A2H A2C A1 A1M A2M A2 A2 A2 A2 A1 A2 A2 A2M A2R A2 A1 A1R A2R A2 A2 Slide 12
Evolution of globin genes 2 Fig. 6.11 Heme Lodish Fig. 3.11 Slide 13
Beta hemoglobin gene cluster Adult: 22(HbA) 22(HbA-2) Fatal: 21 (HbF1) 22 (HbF2) Embryonic: 22 (Hb Gower I) 22 (Hb Gower II) Slide 14
-globin gene cluster in mammals Hoffmann Mol Biol Evol 25:591, 2008 Slide 15
Unprocessed globin pseudogenes What features might a “processed” globin pseudogene have? Slide 16
Dating gene duplication time Fig. 6.12 Fig. 6.9 Slide 17
Dating gene duplication - calculate rates of nt sub (ra and rb) for genes a and b in species 1 and 2 ra = ka/ 2TS rb= kb / 2TS - assume TS is known from geological record • score number of nt sub per site for each gene (that is, a and b) • in the 2 species to determine ka and kb - average rate r = (ra + rb) / 2 • then to estimate TD (where TD = k ab / 2 r) , • need to know kab , the number of • sub per site between genes a and b Slide 18
Getting k: 4 pairwise comparisons 1. Gene a from species 1 and gene b from species 2 2. Gene a from specieis 2 and gene b from species 1 3. Both genes from species 1 4. Both genes from species 2 • depending on degree of divergence may choose to use only • synonymous or only non-synonmyous sites… • if rate constancy holds, the 4 pairwise comparisons should be • approximately equal TD = k ab / 2 r Slide 19
2. Internal domain duplication - repeated sequence may correspond to functional or structural domain within protein - eg. ovomucoid gene in chickens - enzyme which inhibits trypsin and has 3 domains (as a result of duplication events), each of which can bind one molecule of trypsin Slide 20
Ovomucoid gene structure Order of duplication events? Slide 21 Fig. 6.5
Trypsinogen Antifreeze gene in Antarctic cod Fig. 6.6 Slide 22
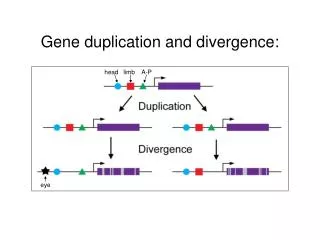
3. Exon/domain shuffling - exon duplication & incorporation into another gene - functional or structural modules form mosaic proteins - may be mediated by intron recombination Gene 1: 1 2 3 4 Duplication of exon 3 & flanking region 3 Gene 2: 3 exon a exon b Slide 23
Phase limitations on exon shuffling If intron lies between 2 codons = “phase 0” If intron between 1st and 2nd nt of codon = “phase 1” If intron between 2nd and 3rd nt of codon = “phase 2” What if the two orange exons were separated by an intron? see Fig. 6.17 Slide 24
Phase limitations on exon shuffling Fig. 6.17 Slide 25
Exons as functional/structural domains Fig. 6.14 F1 = fibronectin module KR = kringle domain Stryer Fig. 10.35 EG = EGF finger module Slide 26
Evolution of new functions • New function through lateral gene transfer • Transformation • Transduction • Conjugation • New function through sequence duplication: • Gene duplication • Internal domain duplication • Exon/domain shuffling • New function without sequence duplication: • Alternative splicing • RNA editing • Overlap genes (alternative transcription) • Different mixtures of gene products How do complex organisms acquire extra genes (for new functions)? … and extra forms of regulation? Slide 27
1. Alternative splicing - single gene can give rise to different mRNA (and different proteins) pre-mRNA mRNA 2 mRNA 1 Slide 28
Possible alternative splicing pathways Fig. 6.21 Example of sex determination pathway in Drosophila see Fig. 6.22 Slide 29
Sex determination in Drosophila Genetics, Vol. 166, 1761-1773 Slide 30
Types of AS in eukaryotes Kim Nucl Acids Res 35: 125, 2006 Slide 31
AS & Mitochondrial gene translocation - organellar genes which move to nucleus during evolution, can only be functional if properly expressed eg. mitochondrial-type rps14 gene located within intron of sdh2 gene SDH2: iron-sulfur subunit of respiratory complex II (succinate dehydrogenase)mt rps14: ribosomal protein S14mt rps14 sdh2 ex1 sdh2 ex2 Figueroa BBRC 271: 380, 2000 Slide 32
2. RNA Editing • Modification of RNA so that message is changed, eg. certain C’s in pre-mRNA changed to U’s. • Occurs in the eukaryotic cell nucleus, cytosol, as well as in mitochondria and plastids, ie. may originate from prokaryotic-like endosymbionts (although not yet observed in prokaryotes) eg. apolipoprotein B in mammals In liver: lipid transport in circulation, LDL receptor binding domain In intestine: truncated protein, role in dietary lipid absorption Lodish Fig. 12-57 Slide 33
RNA editing in T. brucei DNA RNA Fig. 1.7 Slide 34
gRNA and CYb editing Figure 1. Sequence of the 5'CYb mRNA substrates in the editing region (A–C) and full-length gCYb-558 (D). Bold U’s, those added to create partially edited substrates. The gRNA–mRNA anchor is shown for each 5'CYb substrate with the gRNA aligned below the mRNA. The base pairing between the gRNA anchor and the mRNA is shown by Watson–Crick (vertical line) and non-Watson–Crick (colon) base pairs. Changes to the sequence of gCYb-558 are underlined (see Materials and Methods). Leung, S. S., Koslowsky, D. J., 2001. RNA editing in Trypanosoma brucei: characterization of gRNA U-tail interactions with partially edited mRNA substrates. Nucl. Acids Res. 29, 703-709. Slide 35
3. Overlapping genes • DNA region codes for more than one protein • Different reading frames or complementary strand used • In viruses, bacteriophages… (compact genomes) • Rate of evolution expected to be slower for such regions Slide 36
Bacterophage X174 genome Fig. 6.20a Slide 37
Bacterophage X174 genome A protein C protein K protein Fig. 6.20b Slide 38
4. Different mixtures of gene products eg. eye lens crystallin (usually mixture of different crystallins) - in different animals, different proteins have been recruited (eg. LDH, enolase, heat shock proteins…) in response to changing visual environments aquatic – optically dense, high refractive index terrestrial – lens softer, low RI, focus at distance nocturnal vs. diurnal verebrates… Ubiquitous crystallins. -, -, and -crystallins Taxon-specific crystallins: -, -, -, and - crystallins -, -, and -crystallins found in avian and reptilian lenses -, -, and -crystallins found in the lenses of all other vertebrates. Slide 39
Evolution of the lens crystallins r = NADPH-dependent reductase e = lactate dehydrogenase t = enolase Recruitment of various eye lens crystallins during vertebrate evolution Wistow TIBS 1993 Slide 40
Differences in eye lens proteins between octopus & squid Tomarev J Biol Chem 266:24266, 1991 Slide 41
Evolution of eye lens genes 1. Change in regulation so that “housekeeping” gene “up-expressed” in lens multi-functional protein 2. Subsequent aa changes may be favourable for one role, but not other adaptive conflict 3. Resolved by duplication event or reversion back to original function only … and a different gene then recruited for eye lens protein Slide 42