Fatty Acid Catabolism
480 likes | 1.47k Vues
Fatty Acid Catabolism. Lehninger Ch. 17 BIO 322 Recitation 4 / Spring 2013. Role and oxidation of FA. ATP. Electrons from FA to ETC Acetyl CoA may be oxidized completely to in TCA In liver, acetyl CoA ketone bodies

Fatty Acid Catabolism
E N D
Presentation Transcript
Fatty Acid Catabolism Lehninger Ch. 17 BIO 322 Recitation 4 / Spring 2013
Role and oxidation of FA ATP • Electrons from FA to ETC • Acetyl CoA may be oxidized completely to in TCA • In liver, acetyl CoA ketone bodies • Brain and other tissues use as water soluble fuel when glucose is not available. • In higher plants, acetyl CoA is used for biosynthesis, secondary for fuel purposes. • All use 4 step process called, β oxidation, where FA is converted to acetyl CoA. CO2
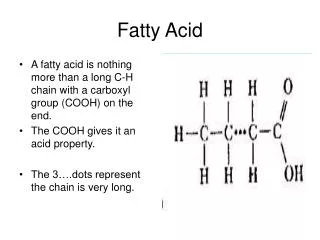
Triacylglycerols • Suitable storage fuels • Long alkyl chains - Complete Oxidation is 38 kJ/g • more than twice of the carbs and proteins. • Insoluble in water, aggregate in lipit droplets • Do not raise the osmolarity of the cytosol. • Inert, no undesired chemical rxn, stored in large quantities. • Ingested TG should be emulsified before digested by water soluble enzymes of the intestine or mobilized via protein attachment. • C-C , C1-coenzyme A, allows oxidation of the fatty acyl groups at C3 or beta position, hence the name beta oxidation
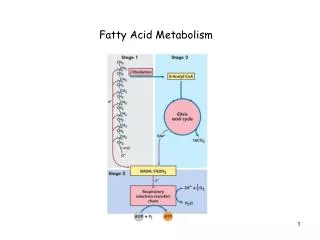
Three Stages The chemical steps of FA oxidation in mitochondria • Oxidation of long chain FA into 2C fragments in the form of acetyl CoA. (β oxidation) (CH 17) • Acetyl CoA – TCA cycle (CH 16) • ETC (CH 19) CO2 H2O
Digestion, Mobilization and Transport of Fats • Fatty acid fuels 3 sources: Diet, Stored as lipid droplets, Synthesis • In industrialized countries ~ 40% daily energy from TG < %30 is recommended. • Provide more than half of the required energy for liver, heart and resting skelatal muscle. • Sole source of energy in hibernating animals and migrating birds Dietary Fats are Absorbed in the Small Intestine • Solubilization of TG into finely dispersed microscopic micelles via bile salts. • Bile Salts • synthesized from cholestrol in liver, stored in gall bladder, released into SI after ingestion of fatty meal. • Biological Detergents, amphipathic • Convert dietary fats into mixed micelles of bile salts and TG – accesible to water soluble lipases in SI
Diffuse into epithelial cells lining the intestinal surface • They are reconverted into TG and packed into lipoprotein aggregates called chylomicrons together with cholestrol and specific proteins. • Apoliproteins (Apo means detached, lipid free form) – lipid binding proteins for transport of TG and other lipid derivatives between organs • VLDL to VHDL (Hydrophobic core, hydrophilic protein side chains and lipid head groups at the surface) • Lipase converts TG into • Diacylglycerols • Monoglycerols • Free FA • Glycerol
Chylomicrons from SI to lymphatic system, then to blood • carried to muscle and adipose tissue via apoCII. • Within the capillaries of the tissues, ApoCII activated lipoprotein lipase hydrolyses TG into FA and glycerol • Taken up by cells • Muscle FA for energy. In adipose FA reesterified for storage as TG. • Chylomicron remnants back to liver. The left over TG by this route may be used for energy or for production of ketone bodies. • More FA than needed, (liver) packed as VLDL – blood to adipose tissue -storage.
Hormones Trigger Mobilization of Stored Triacylglycerols • Neutral lipids are stored in adipocytes, adrenal cortex, ovary, testes – as lipid droplets : Sterol esters and TG in the core surrounded by phospholipids • Coated with Perilipins – restrict acces to the lipid droplet, prevent lipid mobilization. • Hormonal signal for energy (Epi, or glucagon) • Adenylyl cyclase in the plasma memb of adipocyte cAMP cAMP dependent kinase (PKA) phosphorylates perilipin A, phospho-perilipin activated hormone senstive lipase in the cytosol so that hydrolysis of TG into FFA and glycerol can begin. • PKA can also phosphorylate hormone sensitive lipase and duplicate, triplicate its activity. 50X increase via perilipin phosphorylation • Hydrolysis of FA FFA pass from adipocytes into blood • Bind to protein serum albumin ( 10 FA per protein) and carried to the tissue for fuel
Fatty Acids are Activated and Transported into Mitochondria • Enzymes of FA oxidation are located in mitochondrial matrix • Chain length 12 or fewer enter mitoch. w/o any help of a membrane transporter. • 14 or more carbons containing FA, that constitutes most of the FFA from diet cannot pass mitochondrial membrane, • via 3 enzymatic reactions called carnitine shuttle. • Rxn by a family of isozymes (different isozymes specific for short, intermediate, long carbon chains) present in the outer mitoch. membrane, acyl-CoA synthetases • Thioester linkage between carboxyl of FA to thiol of CoA, coupled to ATP cleavage
Fatty acyl-CoAs are high energy compounds – hydrolysis to free FA + acetyl CoA ∆G’º= -31kj/mol • Formation of FA-CoA is made favorable by the hydrolysis of two high energy bonds in ATP. Pyrophosphate hydrolized by inorganic phosphatase • Together with ATP hydrolysis that the overall reaction is pulled towards fatty acyl-CoA formation. • Fatty acyl CoA ester formed in cytosolic side of the outer mitochondrial membrane • Transported either into mitochondrion, oxidized or may be used for membrane lipid synthesis. • If destined to mitochondria • Fatty acyl attached to the hydroxyl group of carnitine to form fatty acyl-carnitine The second reaction of shuttle
Fatty acyl CoA into fatty acyl carnitine via outer mitochondrial membrane enzyme carnitine acyl transferase I. • Either acyl-CoA passes outer membrane and is converted to the carnitine ester in intermembrane space or carnitine ester is formed on the cytosolic face of the outer membrane, then moved across the outer membrane to intermembrane space – current evidence does not reveal which. • Passage into innermemb. space is done via large pores called porin proteins. • Fatty acyl-carnitine ester enter the matrix via faciliated diffusion through acyl carnitine/carnitine transporter of the inner mitochondrial membrane. • At the final, 3rd step, carnitine acyl transferase II regenarates fatty acyl CoA in the matrix, the relased carnitine back to intermemb via same transporter. • Linkage of CoA and fatty acyl CoA pools in the cytosol (Biosynthesis) and mitochondria (Energy).
Oxidation of Fatty Acids • Mitochondrial Oxidation of FA – three stages • First stage – beta oxidation – oxidative removal of two carbon units in the form of acetyl CoA starting from carboxyl end of fatty acyl chain. • Ex. 16 carbon palmitic acid yields 8 two carbon acetyl CoA molecules. • Formation of each acetyl CoA requires removal of 4 hydrogen atoms via dehydrogenases. • Second stage – acetyl CoA into TCA (also in mitochondrial matrix) • Third stage -First two stages produced NADH and FADH2 donate electrons to ETC.
The β oxidation of Saturated Fatty Acids Has Four Basic Steps • First step of fatty acid oxidation – 4 enzymes (Saturated) • Dehydrogenation of fatty-acyl CoA produces a double bond between α and β carbons (C-2,C-3) trans-∆2-enoyl-CoA (delta for position of the double bond, FA nomenclature – HW) • new double bond in trans config, whereas naturally occuring unsaturated fatty acid in cis config. • Acyl-CoA dehydrogenase (three isozymes) : VLCAD – 12 to 18 C, MCAD – 4 to 14 C, SCAD – 4 to 9 C • All three are flavoproteins with FAD, electrons removed from fatty acyl CoA are transferred to FAD and reduced dehydrogenase donates these electrons to electron carrier of ETC flavoproteins (ETF). • Analogous to succinate dehydrogenase rxn, both enzymes are bound to inner membrane of mitochondria, a double bond is introduced into a carboxylic acid α and β carbons , FAD is the electron acceptor, electrons to ETC, 1.5 ATP per electron pair.
Water added to the double bond of trans enoyl CoA to form L stereoisomer of beta-hydroxyacyl-CoA, catalyzed by enoyl-CoA hydratase -- Analogous to fumerase reaction in citric acid cycle. • L- beta hydroxyacyl-CoA is dehydrogenated to form beta-ketoacyl-CoA, by the action of beta-hdroxyacyl CoA dehydrogenase • NAD is the electron acceptor, specific for L stereoisomer • NADH formed donates electrons to NADH dehydrogenase of ETC, analogous to the malate dehyrogenase of citric acid cycle. • Beta-ketoacyl-CoA with free CoA to split off the carboxy terminal 2C fragment of original FA as acetyl CoA by thiolase (acyl-CoA acetyltransferase) • Other product is the coenzyme A thioester of FA, now shortened by 2C. (also called Thiolysis, similar to hydrolysis) • Last 3 steps, fatty acyl chains of 12 or more C are catalyzed by a multienzyme complex in inner mitoch. memb. Trifunctional protein (TFP)
TFP is heterooctamer of α4β4 subunits. • α subunit: 2 activities enoyl-CoA hydratase and β-hydroxyacyl-CoA dehydrogenase • β subunit: thiolase activity • Allows substrate channeling • After TFP shortenes the fatty acyl chain into 12 or fewer carbons, further oxidation is catalyzed by 4 soluble enzymes in the matrix.
Acetyl-CoA Can Be Further Oxidized in the TCA • FADH2 : 1.5 ATP • NADH: 2.5 ATP • In TCA 3NADH, 1 FADH2 , ATP=10 ATP • Beta oxidation + TCA
Oxidation of Unsaturated Fatty Acids • saturated: the SFA’s of a lipid have no double bonds between carbons in chain • polyunsaturated: more than one double bond in the chain • Saturated – single bonds in the carbon chain – typical 4 steps in oxidation • Most of the FA in triacyglycerols and phospholipids are unsaturated – one or more double bonds – cis configuration cannot be acted upon by enoyl-CoA hydratase (acts on trans double bond) • 2 new enzymes: an isomerase for monounsaturated fatty acid and an isomerase together with a reductase for polyunsaturated fatty acids
Odd number FA Coenzyme
FA Oxidation is Tightly Regulated • In liver, fatty acyl-CoA in cytosol • 1) β oxidation in mitochondria • 2) conversion into triacylglycerols and phospholipids in cytosol • Dependent on long chain fatty acyl-CoA transfer into mitoch via carnitine shuttle – rate limiting step of fatty acid oxidation – important point of regulation. Once enter the shuttle committed
When Malonyl-CoA – first intermediate of LCFA sythesis from acetyl-CoA is high • When excess glucose cannot be oxidized or stored as glycogen is converted into FA in cytosol for storage as triacylglycerols • Malonyl CoA inhibits carnitine acytransferase I to stop producing energy from fatty acid oxidation. • NADH/NAD ratio is high β -hydroxyacyl-CoA dehydrogenease is inhibited • High concentrations of acetyl-CoA inhibits thiolase • AMPK also phophorylates ACC (Acetyl-CoA carboxylase) – allows β oxidation
Peroxisomes Also Carry Out β Oxidation • Mitochondria – major site of fatty acid oxidation in animal cells. Peroxisomes - β oxidation in plant cell (similar but not identical) • Intermediates for β acid oxidation of FA are coenzyme A derivaties • Dehydrogenation • Addition of water to the resulting double bond • Oxidation of b-hydroxyacyl CoA to a ketone • Thiolytic clevage by coenzyme A • The difference in the chemistry of the first step • In peroxisomes - Flavoprotein acyl-CoA oxidase that introduces the double bond, passes electrons direct to oxygen, producing hydrogen peroxide (H2O2). • Damaging oxidant cleaved to water and oxygen via catalase • Mitoch – first step electrons – through ETC to O2 for ATP, H2O • Peroxisome – first step electrons – to catalase rxn, which is exothermic, dissipates heat not conserved as ATP.
The ω Oxidation of FA Occurs in the ER • Omega oxidation – minor pathway in mammals, generally active because of a mutation or carnitine deficiency-- defect in β oxidation • Takes place in ER, where carbon most distant to carboxyl group is oxidized. • First step introduces hydroxyl group on the omega carbon, molecular oxygen is used that involves cytochrome p450 and electron donor NADPH, catalyzed by mixed fuction oxidases. • Alcohol dehydrogenase – oxidizes hydroxyl group into an aldehyde • Aldehyde dehydrogenase – oxidizes aldehyde into carboxylic acid, producing a FA with carboxylgroup each end. • Can be attached coenzyme A – mitoch. – beta ox. • Yields dicarboxylic acids such as succinic acid that can enter TCA cycle.
Phytanic Acid Undergoes α Oxidation in Peroxisomes • The presence of methyl group on β carbon of a FA makeβoxidation impossible • These branched FA are catabolized in peroxisomes of animal cells • Example: Phytanic acid, • phytanoyl-CoA is hydroxylated on its alpha carbon that involves the use of molecular oxygen • decarboxlated to form an aldehyde one C shorther • Oxidized to carboxylic acid, that can be further oxided by beta oxidation
Ketone Bodies • Acetyl-CoA formed in the liver • TCA • Ketone bodies (matrix of mitoch.) export to other tissues • Acetone in smaller quantities, is exhaled. • Acetoacetate and d-β-hydroxybutyrate • To other extrahepatic tissues converted to acetyl-CoA for TCA cycle provide energy for heart, muscle and renal cortex. • The production and export of ketone bodies to extrahepatic tissues allows continued oxidation of FA in liver when acetyl-CoA is not being oxidized in TCA cycle.
Ketone Bodies, Formed in Liver, Are Exported to Other Organs as Fuel • Formation of acetoacetate: Enzymatic condensation of two molecules of acetyl-CoA catalyzed by thiolase, reversal of last step of β oxidation. • Acetoacetyl-CoA then condenses with acetyl-CoA to form beta-hydroxy-β-methylglutaryl-CoA (HMG-CoA). • HMG-CoA is cleaved into acetoacetate and acetyl-CoA. • Acetoacetate reduced to d-β-hydroxybutyrate by mitochondrial d-β dehydroxybutyrate dehydrogenease. Specific for d isomer.
Extrahepatic tissues, d-β-hydroxybutyrate oxidized to acetoacetate by d-β hydroxybutyrate dehydrogenase • The acetoacetate is activated to its CoA ester by transfer of CoA from succiniyl CoA of TCA cycle by β-ketoacyl-CoA transferase. • Acetoacetyl-CoA is cleaved into 2 acetyl-CoAs by thiolase. • Used as fuel in extrahepatic tissues, because liver lacks (or little) β-ketoacyl-CoA transferase. • When TCA intermediates are siphoned off for glucose synthesis by gluconeogenesis, cycle slows, so acetyl-CoA oxidation.
Ketone Body Formation and Export from The Liver • Liver contains only a limited amount of CoA, most of it bound to acetyl-CoA, β oxidation slows to supply free coenzyme. • Production and export of ketone bodies frees CoA, allowing continued FA oxidation.