TGN
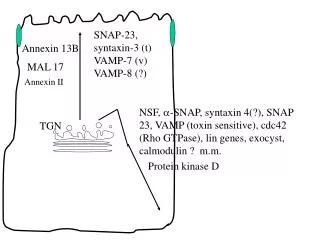
SNAP-23, syntaxin-3 (t) VAMP-7 (v) VAMP-8 (?). Annexin 13B. MAL 17. Annexin II. NSF, -SNAP, syntaxin 4(?), SNAP 23, VAMP (toxin sensitive), cdc42 (Rho GTPase), lin genes, exocyst, calmodulin ? m.m. TGN. Protein kinase D. The major trafficking pathways in polarized epithelial cells.

TGN
E N D
Presentation Transcript
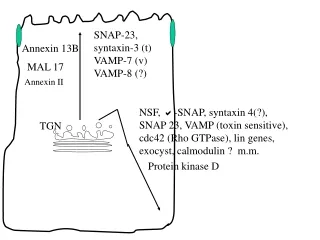
SNAP-23, syntaxin-3 (t) VAMP-7 (v) VAMP-8 (?) Annexin 13B MAL 17 Annexin II NSF, -SNAP, syntaxin 4(?), SNAP 23, VAMP (toxin sensitive), cdc42 (Rho GTPase), lin genes, exocyst, calmodulin ? m.m. TGN Protein kinase D
The major trafficking pathways in polarized epithelial cells.
Mammals Yeast Localisationof differentphosphoinositides, kinasesandphosphatases. If lipid domains are formed withparticular phosphoinositidesearly in the secretory pathway?
Phosphoinositidesare precursors (forløpere) for 2nd messengers and attachment sites for proteins Phosphorylationofphosphatidyl-inositol (PI) givesphosphoinositidesofdifferent kinds. Viktig for transport fra TGN til vakuolen Kinasesthatphosphorylate PI have been found in ER, the nucleus, Golgi, endosomesand the plasma-membrane. The kinases have different specificities – producing different products. LPI: Lysophosphatidylinositol GPI: Glycerophosphatidylinos (Proteinanchorsformed in ER)
*What makes a Golgi-enzymeremain in the Golgiapparatus (longer than proteins in transit)? *What keeps the Golgi cisternaetogether? A typical Golgi glycosyl-transferasespans the membrane once and has the N-terminalend of a short cytoplasmic tail (class II protein) There are hundreds of glycosyltransferases in a mammalian Golgi apparatus.
Sean Munro observed that proteins in the plasmamembrane have a longer hydrofobic transmembrane domain than the Golgienzymes. Impact on localization?
Munro also observedan overrepresentation ofa tyr or trp residueafter the hydrofobic sequence. Impact on localization?
Medial Golgi enzymesfor higher order complexes: Exampel: N-acetylglucosaminyltransferase I (NAGT I) and mannosidase II (Mann II). Trans-Golgi enzymesdo not form such complexes. The luminal domainsare important forcomplex formation, but is the transmembrane domain also important?
Enzymeslate in the Golgi-apparatus havebeen relativelyeasyto study: Example: 2,6-sialyltransferase localizes to the trans-regionof the Golgi by means of the 17-aa transmembrane domain. Enzymesearlier in the Golgi-apparatusthat have been expressed indifferent celllinesoften seemed to localize to the ER. Eventually it became evident that these enzymes needed partner-enzymes to be able to leave the ER in hetero-oligomeric complexes. Example : EXT 1 and EXT 2: Two proteiner that together constitutethe glycosyltransferase activitywhen heparansulfate is polymerised. Must form hetero-oligomeric complexes to leave the ER.
OPEN QUESTIONS: What is the fraction of Golgienzymes passingtheirproperlocalisation before retrieval byvesicles or tubules? Do retrogradely transported vesiclesor tubules bypass several cisterneato fuse with an earlier compartment – only to allow enzymes to move anterogradely towards their proper localisation? Do stable lipid domains form early in the Golgi apparatus (or already in the ER), allowing segregation of apical and basolateral pathways to take place before the trans-Golgi network?
Localisationof proteins to the trans-Golgi network: This compartment is the “exit site” from where transport in several different directions occur. It is probably not avoidable that enzymes acting in this compartment to a large extent are transported to the plasmamembrane. Naturally, endocytosis signals and signals for transport to the TGN have been found in such enzymes (example, the protease furin). Retention of proteins late in the Golgi may be quite complicated.18 different geneshave been identified in yeast (Saccharomyces cerevisiae) wheremutations make the cells incapable of retaining proteins localised late in the Golgiapparatus.
While enzymes in the Golgi apparatus return to the ER upontreatment with Brefeldin A, one class of proteins remains in position:Golgi-MATRIX-proteins.It seems that these proteins govern the appearance and function of the Golgi. ER exit sites, ERGIC and Golgi Cis-Golgi: GRASP 65 + GM 130 + p 115 + rab 1 Medial-Golgi: GRASP 55 + Golgin-45 + rab 2 The Golgi-apparatus collapses without Golgin-45. Golgin-45 does not return to the ER upon BFA-treatment.
Golgi-cisternaeare so close together that it seems as if vesicle budding and fusion only might be feasible at the ends of the cisternea.
Golgi apparatus COP I VTCs COP II Endoplasmic reticulum
COP II vesicle +GTP p115 Rab 1 COP II vesicle