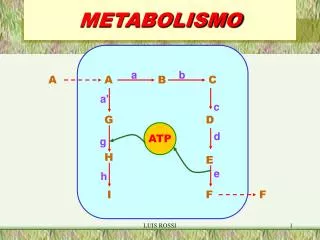
Metabolismo
Metabolismo. Il metabolismo di uno xenobiotico è l’insieme delle trasformazioni chimiche che la molecola subisce all’interno dell’organismo, principalmente ad opera di enzimi .


Metabolismo
E N D
Presentation Transcript
Metabolismo • Il metabolismo di uno xenobiotico è l’insieme delle trasformazioni chimiche che la molecola subisce all’interno dell’organismo, principalmente ad opera di enzimi. • Il metabolismo ha una ‘logica’: trasformare la molecola in modo che possa essere facilmente escreta per via renale o biliare. Dato che le molecole idrofile sono eliminate più rapidamente di quelle lipofile, il metabolismo consiste principalmente nella trasformazione di molecole lipofile in molecole idrofile.
Le sostanze lipofile sono escrete più lentamente di quelle idrofile perché: • sono più facilmente riassorbite nel tubulo renale (o nell’intestino in caso di eliminazione biliare); • si accumulano nel tessuto adiposo
Durante il processo di formazione delle urine, si ha riassorbimento di acqua aumento della concentrazione dei soluti nell’urina gradiente di concentrazione urina-plasma riassorbimento. Le sostanze lipofile sono riassorbite più facilmente perché attraversano meglio lo strato cellulare dell’epitelio tubulare.
E’ stato calcolato che il tempo di dimezzamento (emivita) dell’esobarbitale, che è pari a 5-6 ore, sarebbe, in assenza di trasformazione metabolica, di 2-5 mesi.
Il metabolismo degli xenobiotici è estremamente importante in Tossicologia, poiché può avere sia un effetto protettivo (eliminazione e/o detossificazione dello xenobiotico) sia un effetto dannoso (formazione di metaboliti tossici).
Caratteristiche generali del metabolismo degli xenobiotici • In genere metabolizzano solo gli xenobiotici • Alcuni, tuttavia, sono coinvolti anche nella sintesi o degradazione di composti endogeni
Scarsa selettività per i substrati possibilità di metabolizzare un enorme numero di sostanze con un numero relativamente esiguo di enzimi uno xenobiotico può essere metabolizzato da enzimi diversi lo stesso enzima può catalizzare reazioni diverse sullo stesso substrato in genere uno xenobiotico viene metabolizzato secondo più vie metaboliche
uno stesso enzima può metabolizzare più substrati compresenti nell’organismo possibilità di interazioni (inibizione competitiva, induzione) tra xenobiotici, o tra xenobiotici e composti endogeni
Esempio di interazione: inibizione del metabolismo della terfenadina aritmie Terfenadine (Teldane) Inibitori: succo di pompelmo, ketoconazolo ecc. CYP3A4 Fexofenadine (Telfast)
Una sostanza può inibire (o indurre) una via metabolica di un’altra sostanza, causando una variazione della composizione dei metaboliti
inibita da 2,6-dicloro-4-nitrofenolo aumento tossicità inibito da cimetidina diminuzione tossicità
Variabilità degli enzimi metabolizzanti. 1Variabilità interspecie • In natura, gli xenobiotici con cui vengono in contatto gli organismi animali sono di origine vegetale. • La capacità di diverse specie animali di metabolizzare gli xenobiotici dipende dalla selezione operata dall’esposizione agli xenobiotici vegetali. • Ad es., insetti che si cibano di molte piante diverse possono metabolizzare una varietà di xenobiotici maggiore degli insetti che si alimentano di poche o una sola specie vegetale. I pesci hanno minore capacità metabolizzante dei mammiferi, probabilmente perché possono più facilmente eliminare gli xenobiotici tramite le branchie.
L’evoluzione degli isozimi CYP450 riflette l’esposizione ad un numero sempre maggiore di xenobiotici potenzialmente tossici, sintetizzati dai vegetali a scopo difensivo.
Specie diverse di mammiferi possono avere differenze quali-quantitative della capacità metabolizzante: importanti per l’estrapolazione all’uomo di dati animali. • la maggior parte delle differenze di tossicità tra specie diverse di mammiferi è dovuta a diversità del metabolismo.
Variabilità degli enzimi metabolizzanti. 2Variabilità intraspecie • All’interno di una specie, vi è una variabilità genetica, perché molti enzimi sono polimorfi (più varianti alleliche, con diversa attività; più copie dello stesso gene) differente suscettibilità agli effetti tossici
Variabilità degli enzimi metabolizzanti. 3Variabilità individuale • A livello individuale, la capacità metabolizzante è determinata, oltre che da fattori genetici, anche da fattori endogeni ed ambientali • fattori endogeni: età, sesso, stati fisiologici (ritmi circadiani, ciclo mestruale, gravidanza ecc.), patologie • fattori ambientali: induzione e inibizione enzimatica variazioni della suscettibilità individuale
Gli effetti di un fattore di variabilità dipendono da tutti gli altri fattori. Esempi: • Gli effetti di un inibitore di un enzima saranno più marcati in un individuo con un carenza genetica di quell’enzima. • Le differenze legate al sesso sono molto più forti nel ratto che nell’uomo.
Distribuzione tissutale degli enzimi metabolizzanti • L’organo più ricco di enzimi metabolizzanti è il fegato. • Altri organi o tessuti con significativa capacità metabolica sono i polmoni, i reni, il sangue. • Alcuni tessuti hanno un’elevata concentrazione di enzimi metabolizzanti (mucosa nasale, cristallino), che servono a difendere il tessuto, ma il cui contributo al metabolismo sistemico è pressoché nullo dato il volume del tessuto.
La localizzazione degli enzimi è importante nel determinare l’organo colpito nel caso di formazione di metaboliti tossici. Molti metaboliti tossici sono infatti altamente reattivi e colpiscono quindi solo le cellule dell’organo in cui si formano. Ad es., molti composti sono epatotossici perché i metaboliti reattivi si formano nel fegato. • Inoltre, la tossicità è spesso limitata, all’interno dell’organo, alle cellule che esprimono l’enzima che porta alla formazione del metabolita tossico. Ad es., il paracetamolo e CCl4 causano necrosi centrolobulare perché le cellule della regione centrolobulare sono ricche di citocromo P450.
Localizzazione intracellulare degli enzimi • Molti enzimi hanno diverse isoforme, che hanno spesso diversa localizzazione cellulare. • Una determinata isoforma ha in genere un’unica localizzazione intracellulare. • Nel fegato gli enzimi sono localizzati principalmente nel reticolo endoplasmatico (enzimi microsomiali) e nel citosol. Un numero minore di enzimi è localizzato nei mitocondri, nei lisosomi, nel nucleo.
Fase I: reazioni di ossidazione, riduzione, idrolisi ecc.; funzionalizzazione: introduzione o smascheramento di un gruppo nucleofilo; formazione di elettrofili Fase II: reazioni di coniugazione coniugazione con ac. glucuronico, solfato, acetile, metile coniugazione con glutatione coniugazione con aminoacidi Classificazione delle reazioni di biotrasformazione. Fase I e fase II.
Composti che già contengono un gruppo nucleofilo e composti elettrofili possono essere direttamente coniugati (fase II), ma possono anche subire reazioni di fase I (es. paracetamolo). • Le reazioni di fase II sono in genere più veloci di quelle di fase I basse concentrazioni dei metaboliti di fase I • La capacità delle reazioni di fase I è limitata dalla attività enzimatica (quantità di enzima) • La capacità delle reazioni di fase II è limitata principalmente dalla disponibilità del cofattore
La maggior parte dei metaboliti tossici si forma nelle reazioni di fase I, in particolare nelle reazioni di ossidazione.
Reazioni di fase I • Sono le reazioni di ossidazione, idrolisi, riduzione. • Portano in genere all’introduzione o smascheramento di un gruppo nucleofilo (-OH, -NH2, -SH, -COOH). • Ciò causa solo un modesto aumento dell’idrofilia. Tuttavia, il gruppo funzionale nucleofilo fornisce un punto di attacco per le reazioni di fase II. • Le reazioni di fase I determinano in genere perdita dell’attività farmaco-tossicologica (modificazione della struttura chimica e della capacità di interagire con il recettore). In alcuni casi, tuttavia, i prodotti delle reazioni di fase I sono biologicamente attivi. • Nelle reazioni di fase I, soprattutto le reazioni di ossidazione, si possono formare metaboliti tossici.
Reazioni di ossidazione degli xenobiotici • L’ossidazione è la trasformazione più frequente negli xenobiotici (>75% delle reazioni di fase I) • Le reazioni di ossidazione sono catalizzate principalmente da 7 enzimi, che differiscono per: • i substrati ossidati (ma spesso un substrato può essere ossidato da più enzimi, es. etanolo); • localizzazione tissutale e intracellulare; • attività
Citocromo P450 (CYP 450) • E’ l’enzima più importante di fase I perché metabolizza un gran numero di xenobiotici principale responsabile dell’inattivazione di molti farmaci e tossici diretti. • Coinvolto anche nella biosintesi o degradazione di molti composti endogeni (ormoni steroidei, TXA2, PGI2, vitamine liposolubili, acidi grassi ecc.) • Presente in tutti i tessuti. I livelli più alti sono nel fegato, a livello del reticolo endoplasmatico (frazione microsomiale)
Reazione generale: RH (substrato) + O2 + NADPH + H+ ROH + H2O + NADP+ • E’ una reazione di monoossigenazione in cui un atomo di ossigeno è incorporato nel substrato; l’altro atomo di O è ridotto ad H2O (CYP450 è quindi un’ossidasi a funzione mista), con l’apporto degli equivalenti riducenti (elettroni) provenienti da NADPH. • Il prodotto ROH può subire riarrangiamenti molti prodotti finali, diversi da alcol e fenoli.
CYP450 lega direttamente il substrato e O2. • I 2 elettroni provenienti da NADPH arrivano al complesso CYP450-substrato tramite una flavoproteina, la NADPH-citocromo P450 reduttasi. • Il secondo elettrone può provenire anche dal citocromo b5.
amino ossidasi Reazioni catalizzate dal CYP450: C-ossidazione: idrossilazione alifatica e aromatica. Epossidazione Ossigenazione di un eteroatomo. De-alchilazione di un eteroatomo Trasferimento di un gruppo ossidativo. Scissione di esteri. Deidrogenazione. Riduzione
Ossidazione di composti insaturi: formazione di epossidi • Anelli aromatici e alcheni possono essere ossidati ad epossidi. • L’epossido può isomerizzare a fenolo (aromatici) o a enolo o aldeide (alifatici), non tossici. • Gli epossidi sono in genere (molto) reattivi e si legano ai gruppi nucleofili di proteine e DNA. Es.: bromobenzene, epatotossico; aflatossina B1, cancerogeno. • Gli epossidi di diversi alcheni e alchini, tra cui farmaci come etinilestradiolo, norgestrel, pargilina ecc. sono molto reattivi e si legano al gruppo eme del CYP450, inattivandolo (inibitori suicidi).
I carboni aromatici alogeno-sostituiti non sono ossidati o sono ossidati molto lentamenteresistenza al metabolismo bioaccumulazione (DDT, diossine ecc.)
Gli epossidi di alcheni alogenati sono instabili e per trasposizione formano aldeidi alogenate, che possono essere poi ossidate o ridotte. • La tossicità dipende anche dalla velocità di trasposizione; ad es., il tricloroetene è relativamente poco tossico perché l’epossido traspone velocemente, e l’aldeide viene rapidamente metabolizzata
Ossidazione di idrocarburi policiclici aromatici (IPA) • Dato l’alto numero di posizioni ossidabili, il metabolismo è molto complesso. E’ stato molto studiato perché diversi IPA diventano cancerogeni potenti dopo bioattivazione. • L’ossidazione avviene prevalentemente nelle regioni ad alta densità elettronica (regione L) o in cui si trovano doppi legami con carattere prevalentemente olefinico (regione K). • Sono difficilmente ossidati i legami degli anelli terminali, con carattere prevalentemente aromatico, ed i legami adiacenti ai punti di fusione tra gli anelli.
Spesso si hanno epossidazioni successive in posizioni diverse. • Gli epossidi che si formano nelle regioni ‘Baia’ (Bay region) non sono attaccabili dall’epossido idrolasi e dalla glutatione-S-transferasi cancerogenesi (es., benzo[a]pirene)
Ossidazione di eteroatomi da parte di CYP450 Composti solforati • Mercaptani alcoli • Tioeteri solfossidi ( solfoni) • Solfossidi solfoni Gli intermedi R-S-OH (acidi solfonici) sono reattivi, potenzialmente tossici Reazioni catalizzate anche dalle monoossigenasi flaviniche (FMO)
N-ossidazione Reazioni catalizzate da CYP450 e/o FMO • Formazione di N-ossidi. P450: N ricchi di elettroni (piridine, chinoline); FMO: N poveri di elettroni (amine terziarie) • Formazione di N-idrossilamine: amine aromatiche primarie e secondarie. Le N-idrossilamine possono essere O-coniugate con gruppi acetile o solfato uscita di di gruppi acetossi o solfato ioni nitrenio reattivi metaboliti cancerogeni. Le N-idrossilamine aromatiche possono anche essere ossidate a nitroso- e nitro-composti
N-dealchilazione (amine alifatiche primarie e secondarie): catalizzata solo da CYP450. • In generale, CYP450 catalizza la N-dealchilazione perché l’intermedio è un radicale cationico instabile perdita di un protone in alfa idrossilazione del C-alfa dealchilazione (formazione dell’aldeide) • Ovviamente questo meccanismo non avviene se non ci sono protoni in alfa (amine terziarie, aromatiche primarie) o se il radicale cationico è stabilizzato da gruppi elettron-donatori.
La dealchilazione di metil-amine porta alla formazione di formaldeide acido formico CO2. • Misurando la quantità di 13CO2 dopo somministrazione di un amina marcata con 13C sul metile (es. eritromicina) si può misurare l’attività di CYP450 in vivo.
Dealchilazione di nitrosoamine: porta alla formazione di un diazoidrossido diazoione catione alchilico cancerogenesi. La formazione di nitrosoamine può avvenire in vivo, a partire da amine secondarie e nitriti, catalizzata dal pH acido dello stomaco. • L’ossidazione delle idrazine forma composti instabili che decompongono con formazione di cationi (o radicali) alchilici cancerogenesi (1,2 dimetilidrazina), epatotossicità (isoniazide)
Dealogenazione di idrocarburi alogenati • Dealogenazione ossidativa, seguita da eliminazione di HX, con formazione di alogenuri acilici o aldeidi • Dealogenazione riduttiva: sostituzione dell’atomo di alogeno con H. Anche la riduzione è catalizzata da CYP450, quando la tensione di O2 è bassa. All’aumentare dei sostituenti alogenati aumenta il potenziale di ossidazione preferenzialmente riduzione • Rimozione di due atomi di alogeno adiacenti alchene
Metaboliti tossici delle reazioni di dealogenazione • La dealogenazione riduttiva di CCl4 ed altri idrocarburi alogenati ha come intermedio il radicale CCl3. perossidazione lipidica • Si formano inoltre altri metaboliti tossici, come il fosgene tossicità epatica e renale Nella terapia dell’avvelenamento da CCl4 si somministra O2 per inibirne la riduzione.
Alotano • Dealogenazione riduttiva radicale perossidazione epatotossicità diretta, con severità specie-dipendente (severa nel ratto, lieve nella cavia) • Dealogenazione ossidativa formazione di cloruro di trifluoroacetile addotti proteici formazione di neoantigeni epatite immuno-mediata nell’uomo (e nella cavia). • Rara, si manifesta dopo esposizione ripetuta. Sensibilizzazione crociata con enflurano e forse altri anestetici alogenati l’enflurano causa epatotossicità in soggetti precedentemente esposti ad alotano