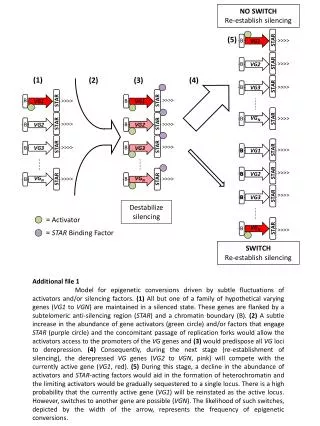
RNAi and chromatin silencing
250 likes | 414 Vues
RNAi and chromatin silencing. Heterochromatin - condensed regions during cell cycle (E. Heitz,1928) - transcriptionally repressed and highly condensed structure - recombination suppression, chromosome segregation, gene silencing,

RNAi and chromatin silencing
E N D
Presentation Transcript
Heterochromatin - condensed regions during cell cycle (E. Heitz,1928) - transcriptionally repressed and highly condensed structure - recombination suppression, chromosome segregation, gene silencing, - DNA methylation and histone modifications control diverse chromatin states and architectures. Recent work indicates that this highly condensed chromosomal material is transcribed, and rapidly silenced, by an orchestrated sequence of events directed by RNA interference (RNAi). transcription and noncoding RNAs
convergent view of the molecular mechanisms that underlie heterochromatin assembly - centromeric heterochromatin formation in fission yeast - RNA-directed DNA methylation in plants - X chromosome inactivation (XCI) and imprinting in mammals.
Science297, 1833-1837, 2002 Regulation of HeterochromaticSilencing and Histone H3 Lysine-9 Methylation by RNAi Thomas A. Volpe,Catherine Kidner, Ira M. Hall, Grace Teng, Shiv I. S. Grewal, Robert A. Martienssen1 Eukaryoticheterochromatinischaracterized by a high density of repeats and transposons, aswellas by modifiedhistones, and influencesboth gene expression and chromosomesegregation. In the fissionyeastSchizosaccharomycespombe, wedeleted the argonaute, dicer, and RNA-dependent RNA polymerase gene homologs, whichencode part of the machineryresponsible for RNA interference (RNAi). Deletionresults in the aberrantaccumulation of complementarytranscripts from centromericheterochromaticrepeats. Thisisaccompanied by transcriptional de-repressionof transgenesintegratedat the centromere, loss of histone H3 lysine-9 methylation, and impairment of centromerefunction. We propose that double-stranded RNA arising from centromericrepeats targets formation and maintenance of heterochromatinthroughRNAi.
Centromeric silencing is relieved in ago1-, dcr1-, and rdp1- mutant strains compared to wild type. Diagram of the three S. pombe centromeres (A) including locations of ura4+ transgenes as well as outermost (otr), innermost (imr), and central (cnt) centromeric regions. Conserved dg (green) and dh (red) repeats are indicated as arrows. Regions containing one or more tRNA genes are indicated by yellow boxes. Northern analysis (B) of RNA transcripts transcribed from centromeric ura4+ transgenes and a ura4+ (DS/E) mini-gene located on the chromosome arm. Transcripts derived from centromeric repeats were detected by Northern blotting (C and D) using probes specific for dg centromeric repeats
The RNAi machinery is required for the initiation and maintenance of the heterochromatic state of centromeric repeats. Reverse strand centromeric transcription occurs in wild-type cells and is degraded posttranscriptionally by the RNAi machinery. Low-level transcription from the forward strand and/or amplification by Rdp1 results in generation of dsRNA, which is converted to siRNA by RNAi. Rdp1, bound to the chromatin, promotes targeting of histone modifications to specific sequences via siRNA, resulting in maintenance of the heterochromatic state (HMT, histone methyl transferase).
Modifications of histone H3. Lysine residues on histone H3 can be mono-, di- or tri-methylated. Shown are modifications H3K4me1, H3K4me3 and H3K36me3, which mark active/poised enhancers, active/poised promoters and actively transcribed regions, respectively. me, methylation. At least six mammalian homologs of COMPASS exist, including MLL1–4 complexes, hSET1A and hSET1B, and their recruitment to active promoters can result in H3K4me3
nuc nuc nuc markers of heterochromatin in mosteukaryotes • histone H3 methylatedatlysine 9 (H3K9me) • Su(VAR)2-5 - HP1- (Swi6 in S.pombe) binds H3K9me • SU(VAR)3-9 -histone H3-K9 methyltransferaseHMTase(Clr4in S.pombe - Ezh2 and G9A in mammals) clr4 H3K9me swi6 swi6 swi6 nuc nuc nuc Propagation of heterochromatin platform for the recruitment of other repression activities
Transcriptional regulation through H3K27 methylation. (A) Transcriptional repression regulated by the polycomb repressive complex 2 (PRC2) including EZH1 or EZH2. PRC2 complexes, which include EED, SUZ12, RbAP46/48 and EZH2, catalyze H3K27 di- and tri-methylation. This in turn leads to a more condensed chromatin state and transcriptional repression. PRC2 complexes containing EZH1 can also catalyze H3K27 methylation, or compact chromatin through other mechanism. G9a has H3K27 methyltransferase activity and also affects gene silencing. (B) Transcriptional activation regulated by UTX and JMJD3. UTX and JMJD3 are demethylases that form complexes with MLL, RbBP5 and WDR5. The removal of methyl groups from H3K27me3 leads to transcriptionally active
Active genes are frequently marked by H3K4me3 (see page 407); this modification is presumably removed by LSD1 (which has not yet been characterized in Drosophila). H3K9 is normally acetylated in euchromatin, and this modification must be removed by a histone deacetylase, typically HDAC1. Phosphorylation of H3S10 can interfere with the methylation of H3K9; its dephosphorylation might involve a phosphatase targeted through the carboxy terminus of the protein kinase JIL1 (ref. 10). These transitions set the stage for acquisition of the modifications that are associated with silencing: these include the methylation of H3K9 by SU(VAR)3-9 or another histone methyltransferase, the binding of HP1, and the subsequent methylation of H4K20 by SUV4-20 (an enzyme that is recruited by HP1). Other silencing marks such as methylation of H3K27 by E(Z) (enhancer of zeste; not shown) seem to be relevant in some regions, although this mark is more prominently used by the Polycomb system. Supporting data come from genetic identification of modifiers of PEV, as well as biochemical characterization of the activities of such modifiers and tests of protein–protein interactions10. (Figure adapted, with permission, from ref. 10.)
Chromatin silencing RITS nuc nuc nuc S.pombe DCR1 dsRNA dsRNA synthesis Repeats and transposons siRNA RDRC RITS nuc nuc nuc RITS active RITS RDRC recruitment AMPLIFICATION Clr4 Methyl transf clr4 RITS recruitment RITS swi6 swi6 swi6 H3K9me nuc nuc nuc Swi6-dependent chromatin formation
RNAi factors such as RITS, RDRC and Dicer, involved in processing of repeat transcripts (red line) into siRNAs, are required for targeting of ClrC to the heterochromatic repeats. siRNA-bound Ago1 is likely to specify the targeting of RITS to nascent repeat transcripts. RITS then facilitates ClrC loading. Rik1 might also directly associate with the repeat transcript and/or some part of the elongating RNAPII complex, thus promoting ClrC loading to nucleate heterochromatin. After the initial methylation of H3K9 by ClrC, Clr4 bound to H3K9me could modify adjacent nucleosomes creating additional binding sites for ClrC and other chromodomain proteins including Swi6 and Chp2 (HPs), which in turn mediate recruitment of factors such as SHREC, thereby promoting higher-order chromatin organization. Swi6 could further contribute to long-range heterochromatin spreading by promoting higher-order chromatin organization by forming oligomers and stabilizing the ClrC binding to chromatin. Boundary DNA elements block inappropriate spreading of heterochromatin into euchromatic regions. Green flag, histone acetylation; red lollipops, H3K9me. RNA dependent RNA polymerase complex - RDRC promote double-stranded RNA (dsRNA) synthesis
Marc Buhler – Basel Nat Struct Mol Biol. 2013 Aug;20(8):994-1000 ncRNA can act as EVICTORS ncRNAs can counteract the spreading of heterochromatin into neighboring euchromatin BORDERLINE prevents spreading of the HP1 protein Swi6 and histone H3 Lys9 methylation beyond the pericentromeric repeat region of S.pombe It acts in a locus-dependent manner and isprocessed by Dicer into sRNAs It evicts HP1 from the chromatin Borderline acts as an allosteric regulator – Mass spec diff. interctors if SWI-6 alone or bound to the ncRNA
Role of non coding RNAs and RNAi in DNA methylation and histone modification
Role of non coding RNAs and RNAi in DNA methylation and histone modification In plants two different RNA polymerase (IV and V) are required for heterochromatin assembly and for transposon silencing Pol IV transcription of target loci with RDR2 and DCL3 24 nt long siRNA AGO4 DRM2 and Pol V + DNA methylation RNA dep-RNA pol dicer methyltransferase
Pol IVa and Pol IVb are players in the RdDM pathway RNA polymerase IV silences certain transposonsand repetitive DNA in a short interfering RNA pathway involvingRNA-dependent RNA polymerase 2 and Dicer-like 3 The existence of this distinct silencing polymerase may explain the paradoxical involvement of an RNA silencing pathway in maintenance of transcriptional silencing. higher plants have five multi-subunit nuclear RNA polymerases: the ubiquitous Pol I, II and III, which are essential for viability; plus two non-essential polymerases, Pol IVa and Pol Ivb which specialize in small RNA-mediated gene silencing pathways In the RNA-directed DNA methylation (RdDM) pathway of transcriptional gene silencing double-stranded RNAs generated with the involvement of RDR2 are cleaved by DCL3, and the resulting siRNAs are loaded into AGO4–RISC and/or AGO6–RISC complexes that mediate the de novo methylation of cytosines within DNA sequences complementary to the siRNAs
RNA silencing and DNA methylation in Arabidopsis • Cytosine DNA methylation silences harmful DNAs such as transposons and retroviruses • Maintenance DNA methyltransferases propagate pre-existing DNA methylation in the CG sequence context by methylating hemi-methylated sites after DNA replication • RNA Silencing Genes Control de Novo DNA Methylation in Arabidopsis
RNAi participates in gene regulation also in euchromatin HP1 recruits cohesin the protein complex responsible for sister chromatid cohesion during cell division cohesin is believed to promote transcription termination between convergent genes - cohesin Gullerova M, Proudfoot NJ.Cell 2008 132:983-95.
Figura 3 Modificazione della cromatina rasiRNA RITS nuc nuc Taglio dell’RNA 7mG siRNA miRNA AAA RISC P Repressione traduzionale miRNA miRNP AAA 7mG
RNA silencing pathways in different organisms. Long dsRNA and miRNA precursors are processed to siRNA/miRNA duplexes by the RNase-III-like enzyme Dicer. The short dsRNAs are subsequently unwound and assembled into effector complexes: RISC, RITS (RNA-induced transcriptional silencing) or miRNP. RISC mediates mRNA-target degradation, miRNPs guide translational repression of target mRNAs, and the RITS complex guides the condensation of heterochromatin. In animals, siRNAs guide cleavage of complementary target RNAs, whereas miRNAs mediate translational repression of mRNA targets. rasiRNAs guide chromatin modification. S. pombe, C. elegans and mammals carry only one Dicer gene. In D. melanogaster and A. thaliana, specialized Dicer or DLC proteins preferentially process long dsRNA or miRNA precursors. 7mG, 7-methyl guanine; AAAA, poly-adenosine tail; Me, methyl group; P, 5' phosphate.


![RNA interference (RNAi) [aka post-transcriptional gene silencing (PTGS)]](https://cdn4.slideserve.com/584681/rna-interference-rnai-aka-post-transcriptional-dt.jpg)