Download

1 / 64
640 likes | 663 Vues
This overview explores the reverse-engineering of transcriptional networks, focusing on inferring regulatory mechanisms from gene expression data and identifying co-expressed genes using microarray technology. It delves into the analysis of cis-regulatory elements and transcriptional modules in yeast and human cell cycles, along with the comparative genomics approach to conserve regulatory elements and detect novel targets. Additionally, the regulation of gene expression by microRNAs and the identification of putative miRNA targets are discussed, showcasing how computational tools and comparative analysis aid in understanding complex transcriptional networks.

E N D
Overview:Functional Genomics Dissections of Transcriptional Networks Rani Elkon Ron Shamir, Yossi Shiloh
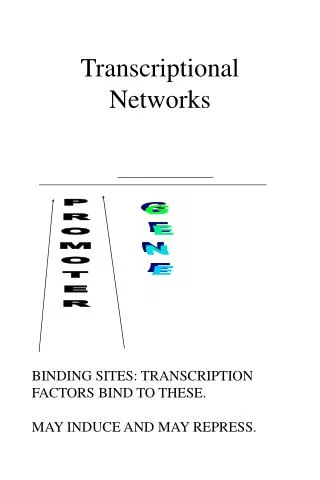
‘Reverse engineering’ of transcriptional networks • Infers regulatory mechanisms from gene expression data • Assumption: co-expression → transcriptional co-regulation → common cis-regulatory promoter elements • Step 1: Identification of co-expressed genes using microarray technology (clustering algs) • Step 2: Computational identification of cis-regulatory elements that are over-represented in promoters of the co-expressed genes • Such methodologies were first demonstrated in yeast
Reverse-engineering of the Yeast Cell-Cycle • Expression profiles were recorded in synchronized yeast cells in 10 min intervals over 2 cell cycles. • ~ 500 ORFs showed a periodic expression pattern
Reverse-engineering of the Human Cell-Cycle • Whitfield et al. recorded expression profiles during the progression of human cell cycle. • 874 genes showed periodic expression patterns, and were partitioned into five clusters (G1/S, S, G2, G2/M and M/G1). • We applied promoter analysis to these 5 clusters
p = 1.2x10-8 (true positive) 78 promoters (92 hits) p = 1.2x10-11 (152, 203) p = 8x10-4 (20, 25)
Transcriptional Modules I: Co-occurrence • Transcriptional regulation is combinatorial Promoter #1 Promoter #n
Defining transcriptional modules: • Co-occurrence • Positional bias (distance) • Orientational bias (order)
Conservation of Regulatory Elements Gene “DNA replication licensing factor MCM6”: (G1/S)
Human Cell Cycle Revisited • We detected global enrichments that pointed to major TFs in human cell cycle regulation. • However, we did not report on specific target genes due to high rate of false positive hits. • Comparative Genomics greatly boosts the specificity of in-silico detection of regulatory elements. • It now allows us to pinpoint TF targets with high confidence.
E2F Human-Mouse Conserved Hits 16,299 human-mouse ortholog promoters (Ensembl)
CHR Regulatory Element • Cell-cycle Homology Region • To date, CHR was experimentally identified on 7 cell cycle-regulated promoters: • including CDC2, CCNB1,CCNB2 and CDC25C (major regulators of G2-M)
Transcriptional Modules Promoter #1 CHR and NF-Y elements show significant co-occurrence rate (p<10-11) Promoter #n
CHR-NF-Y Module 16,299 Hs-Mm ortholog promoters; NFY-CHR putative targets: 71 CHR-NFY: novel transcriptional module with a pivotal role in G2-M regulation
G2/M G1/S CHR-NFY Module Dictates Expression that is Specific to G2/M
CHR-NFY Module – False Positive Rate Comparative genomics yields highly specific identification of novel CHR-NFY cell-cycle targets
Regulation of CyclinB-CDC2 activity Rho GTPases pathways Cytokinesis Regulation of the mitotic spindle assembly Regulation of the kinetochore apparatus NovelCHR-NFY Targets in the G2-M Network
Mature miRNA (~ 22 bp) tend to: • Start with a “U” base • Bind their target mRNAs at sites of length 8 bp. • Target site is complementary to positions 1-8 of the mature miRNA. • Assumed to play major regulatory function during development (many show tissue-specific expression pattern)
Transfected two miRNAs into Hela human cells and examined changes in mRNA expression profiles: • miR-1: expressed in skeletal muscle • miR-124: expressed in brain • 96 and 174 genes were significantly down-regulated by miR-1 and miR-124, respectively • Comparison with human tissue expression atlas: • Genes down-regulated by miR-1 are expressed at lower levels in skeletal muscle and heart than in other tissues • Genes down-regulated by miR-124 are expressed at lower levels in the brain than in other tissues • Searching for enriched signals in the 3’-UTRs of the down-regulated genes discovered the cognate binding sites
Computational identification of putative miRNA targets – scan 3’-UTRs for putative target sites • Anti-Correlation between the expression pattern of miRs and their putative targets (using the human tissue gene expression atlas) • Genes expressed at the same time and place as a miRNA evolved to avoid sites matching the miRNA
Comparative analysis of promoter and 3’-UTR regulatory motifs using the human, mouse, rat and dog genomes. • Search for highly conserved motifs (degenerate strings, 6-18 bases) • Motif Conservation Score (MCS): Z score of the proportion of the conserved occurrences of a motif relative to the conservation rate of comparable random motifs. • Promoters (-2 kb to + 2kb relative TSS): • 174 highly conserved motifs (MCS > 6): • 69 – known (out of 123 TRANSFAC motifs, 56%) • 105 potentially novel regulatory elements
Demonstrating biological function for the discovered motifs: • Correlate the occurrence of a motif with tissue-specific gene expression (using data from the human tissue expression atlas) • Target sets of 86% (59 out of 69) of the known motifs showed significant tissue-specific expression • 53 out of the 105 (50%) novel motifs • Examine positional bias of the motif hits
3’ UTR signals: • 106 highly conserved motifs (MCS > 6) • Hypothesis: function as binding sites for miRNAs • Many of the discovered motifs show features of miRNA binding sites: • Strong strand biasof the conservation rate • consistent with a role in post-transcriptional regulation, acting at the RNA rather than DNA level • Biased length distribution: strong peak at 8 bp • High rate of “A” in position #8 • Search for matches of the 8-mer motifs to the known human miRs: • in 95% of the cases the matches begins at position 1 or 2 of the mature miRNAs.
Systems-level analysis of the DNA damage response in yeast by an integrated approach that combines: • Genome-wide profiling of TF-promoter binding (ChIP-chip data) • Expression profiling (in deleted and w.t. strains) • Phenotyping sensitivity to DNA damage in deleted strains • Wide scale protein-protein interaction data
Systematic screen for TFs involved in the DNA damage response: • 30 (out of 141) TFs based on either: • Expression: differentially expressed after DNA damage • Binding: bind promoters of genes induced by DNA damage • Sensitivity: TF-mutant strain is hyper-sensitive to DNA-damaging agent • TF-promoter binding profiling (Chip-chip) for each of these 30 TFs, without and after exposure to DNA damaging agent
Validation of functional roles of the measured TF-promoter binding interactions: • Gene expression profiling in w.t. and deleted strains (27 out of the 30 are non-essential) without and after exposure to DNA damaging agent • “Deletion Buffering”: genes that respond to the damage in w.t. but become unresponsive in a specific TF-deleted strain • Only 11% (37 out of 341) of the observed deletion-buffering events could be explained by direct TF-promoter interaction • The rest are probably mediated by longer, indirect, regulatory pathwayslinking the deleted TF and the buffered gene
Physical pathways that explain indirect deletion-buffering events were searched for using Bayesian modeling procedure • Utilized various data sources: • TF-promoter binding data measured in this study • Tf-promoter binding data measured for all yeast TFs (in nominal conditions) • 14K high-throughput protein-protein interactions (in nominal conditions) • The inferred network explains a total of 82 deletion-buffering events.
and in human cells? Small-interfering RNA (siRNA)
RNA Interference (RNAi) • A major technological breakthrough in biomedical research • Allows rapid establishment of mammalian cell lines which are stably knocked-down for any gene of interest – pivotal tool in functional genomics • Efforts to establish cell lines in which specific genes are silenced, eventually spanning most of the genome
The combination of RNAi and microarrays holds promise as a powerful tool for a systematic, genome-wide, dissection of transcriptional networks in human cells
Experiment Goal • Proof of principle that RNAi+microarrays can "deliver" • Focus on transcriptional network induced by DNA damage as a test case
Transcriptional network induced by DNA double strand breaks ATM DNA Double Strand Breaks AP-1 CREB NF-kB E2F1 p53 g13 g12 g11 g10 g9 g8 g7 g6 g5 g4 g3 g2 g1
Heatmap colors: Red – above average induction Black – average induction Green – below average induction • 26 genes whose activation is: • Strongly reduced in the absence of ATM and Rel-A • Partially reduced in the absence of p53 • ATM-NFκB-dependent cluster, partial role for p53
46 genes whose activation is: • Strongly attenuated in the absence of ATM and p53 • Not affected by the absence of Rel-A • ATM-p53-dependent cluster
Response of known NF-κB targets • Knocking down Rel-A subunit of NF-κB abolished the induction of known NF-κB targets • ATM is required for the activation of the NF-κB mediated transcriptional response • p53 plays a positive role in the activation of NF-κB targets (?)
Response of known p53 targets • Knocking down p53 attenuated the induction of its known targets • ATM is required for the activation of p53 targets • NF-κB plays an inhibitory role in the induction of some components in the p53 pathway (?)