Download
1 / 28
570 likes | 2.57k Vues
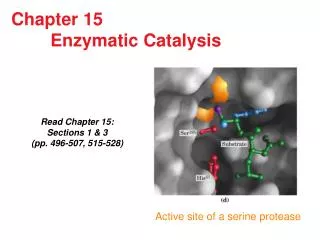
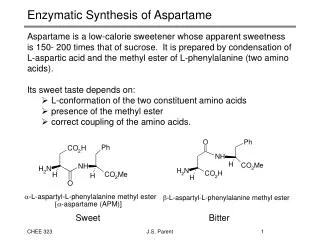
Enzymatic Synthesis of Aspartame. Aspartame is a low-calorie sweetener whose apparent sweetness is 150- 200 times that of sucrose. It is prepared by condensation of L-aspartic acid and the methyl ester of L-phenylalanine (two amino acids). Its sweet taste depends on:

E N D
Enzymatic Synthesis of Aspartame • Aspartame is a low-calorie sweetener whose apparent sweetness is 150- 200 times that of sucrose. It is prepared by condensation of L-aspartic acid and the methyl ester of L-phenylalanine (two amino acids). • Its sweet taste depends on: • L-conformation of the two constituent amino acids • presence of the methyl ester • correct coupling of the amino acids. • Sweet Bitter J.S. Parent
Industrial Enzymatic Synthesis of Aspartame • The unique regio and stereoselectivity afforded by enzymes has been exploited on an industrial scale Aspartame production. • The process employs a protease, • thermolysin, to catalyze the • condensation of the modified Asp • and Phe). • The forward reaction is written as: • Note however, that the synthesis reaction is equilibrium limited by the reverse (hydrolysis) reaction for which proteases are known. Furthermore, the equilibrium strongly favours hydrolysis. J.S. Parent
Structural Properties of Thermolysin • Thermolysin is a metalloenzyme (316 amino acids) requiring a zinc ion and four calcium ions to maintain an active tertiary structure. • Two distinct hemispheres exist with a zinc atom located at the bottom of the cleft. Three residues (142, 146 and 166) serve as ligands for zinc. Calcium is a structural element, and is not believed to interact with the substrate at the active site. • Open circles: a-carbon positions Stippled circle: zinc with its three protein ligands as broken lines • Solid circles: four calcium atoms J.S. Parent
Chemical Properties of Thermolysin • Thermolysin is an extracellular enzyme produced by a bacterial strain that can withstand high temperatures. Hence, themolysin has a temperature stability that is superiour to most enzymes. • Thermolysin is classified as a protease, in that it catalyzes the cleavage of the peptide bonds that constitute proteins. • The term endopeptidase applies, as the internal bonds in polypeptides are susceptible to the action of thermolysin • The term neutral protease applies, as the pH optimum lies about pH 7.5 • The term metalloenzyme is appropriate, given the necessity of zinc at the active site and the requirement for calcium to maintain an active tertiary structure. Chelating agents deactivate thermolysin. • Enzymes of this class demonstrate substrate specificity which requires a hydrophobic amino acid such as phenylalanine as the residue whose amido group is cleaved. J.S. Parent
Kinetics of the Aspartame Synthesis • The rate of APM production is first-order with respect to the total concentration of enzyme [Eo], and a bell-shaped pH-rate profile with the highest activity at pH 7.5 is observed. • Shown is a typical time course of the thermolysin catalysed condensation of • N-benzyloxycarbonylaspartic acid with phenylalanine methyl ester. Initial rate measurements (from t=0 to t=10 min) as a function of reagent concentrations define the overall reaction kinetics. • [Z-L-Asp] = 1.82 x 10-2 M • [L-PM] = 3.64 x 10-2 M • [Eo] = 4.85 x I0-6 M • pH = 6.5; m = 0.364 M • T = 40C J.S. Parent
Influence of [PM] on the Condensation Rate APM synthesis is first-order WRT phenylalananine methyl ester, with no apparent saturation behaviour that is common in enzyme-mediated reactions. Note that the presence of D-PM has no effect on the reaction rate, and it is not found in the product. * [L-PM] = 1.82x10-2 M with [D-PM] = 9.09x10-3 M ** [L-PM ] = 3.64 x10-2 M with [D-PM] = 1.82 x10-2 M [Z-L-Asp] = 1.82 x 10-2 M [Eo] = 4.85 x I0-6 M pH 6.5; m 0.364 M; 40C L-PM D,L-PM J.S. Parent
Influence of [Z-L-Asp] on the Condensation Rate A plot of [Z-L-Asp] against the APM production rate shows saturation of the rate, typical Michaelis-Menten behaviour. Rate retardation occurs in the presence of Z-D-Asp, indicating that the enantiomer acts as a competitive inhibitor. Hence only pure L-Asp can be used in APM synthesis, while racemic mixtures of D,L-PM can be accommodated. [L-PM] = 3.64 x 10-2 M [Eo] = 4.85 x I0-6 M pH 6.5; m 0.364 M; 40C Pure Z-L-Asp 9.1x10-3 M Z-D-Asp added J.S. Parent
Proposed Reaction Mechanism • Competitive inhibitors reduce the rate of product formation through binding the enzyme in an inactive form. • Often these inhibitors are structurally similar to the substrate, and therefore are capable of binding at the active site • Enzyme-bound inhibitor either lacks a needed functional group or is held in an unsuitable position for reaction. • We have seen an example of this behaviour in aspartame production, where the enantiomer of L-Asp inhibited the reaction. A plausible mechanism for this inhibition is shown below: • Note that Z-D-Asp binds thermolysin in an • inactive state, thereby reducing the active • enzyme concentration and lowering the • reaction rate. J.S. Parent
Competitive Inhibition by Z-D-Asp • From this proposed mechanism we can derive a rate expression that accounts for competitive inhibition. • r1: • r3: • r2: • Assigning r2 as the rate determining step of the process, we find the reaction velocity is: J.S. Parent
Validating the Proposed Reaction Scheme • Although more sophisticated regression techniques are available, the simplest means of testing the model is to linearize the rate expression • by inverting it: • A plot of 1/rate versus 1/[Z-L-Asp] should be linear, with a slope of K1(1+[Z-DAsp]/K3)/(k2[E]T[L-PM]) and an intercept 1/(k2[E]T[L-PM]) • This is commonly referred to as a Lineweaver-Burk plot • It is necessary that the data fit the rate expression, but it is not sufficient proof that the mechanism is correct • From the slope, intercept, [E]T and [L-PM], numerical estimates of K1 and k2 can be derived. J.S. Parent
Lineweaver-Burk Plot of the Kinetic Data • Plotting the inverse of the APM production rate (moleL-1s-1) against 1/[Z-L-Asp] reveals a linear relationship • The proposed mechanism is consistent with the kinetic data, and may be correct. • From the slopes and intercepts, k2 = 2.65 L mole-1 s-1 K1 = 1.03x10-2 mole L-1 • K3 = 2.35x10-2 mole L-1 • Line A: no Z-D-Asp; • Line B: [Z-D-Asp]=9.09x10-3 M • [L-PM] = 1.82 x 10-2 M • [Eo] = 4.85 x 10-6 M • pH 6.5; m 0.364 M; 40C J.S. Parent
Isolation of the Aspartame Product • Proteases are recognized as catalysts for peptide bond cleavage, and using them to catalyze the reverse condensation reaction can be problematic. • The equilibrium constant derived from • the Gibbs energies of the reaction • components is quite small, making the • conversion of a standard batch reaction • equilibrium limited. J.S. Parent
Isolation of the Aspartame Product • Luckily APM forms, via its free side-chain carboxylic acid, a sparingly soluble addition compound with excess PM. • The synthesis can be driven using LeChatalier’s Principle by removal of the precipitation of the product. • Once isolated from the enzyme, hydrolysis of Z-APM is no longer a concern, excess PM can be removed and the product can be deprotected to yield aspartame. J.S. Parent
Effects of Immobilization on Enzyme Stability and Use • Design of enzymatic processes requires knowledge of: • reactant and product selectivity • thermodynamic equilibria that may limit product yield • reaction rate as a function of process conditions ([Enzyme], [substrate(s)], [Inhibitors], temperature, pH, …) • Two design issues that we have not considered are: • enzyme stability • efficiency losses associated with the use of homogeneous (soluble) catalysts • Immobilization of an enzyme allows • it to be retained in a continuous reactor, • but its initial activity and its stability • directly influence its usefulness • in industrial applications. J.S. Parent
Enzyme Stability • Although enzyme storage stability is important, it is the operational stability of an enzyme that governs its reactor performance. • Operation stability is a complex function of temperature, pH, [substrate] and the presence of destabilizing agents. • Generally, the rate of free enzyme deactivation is first order with a deactivation constant, kd: • Integrating this expression • yields the concentration of • active enzyme as a function • of time: J.S. Parent
Effect of Thermolysin Instability on APM Production • Recall the rate expression developed for APM synthesis by thermolysin: • If thermolysin deactivation were adequately described as a first order process, the observed reaction rate would have an explicit time dependence, as shown below: • where [E]T,o represents the initial enzyme concentration and kd is the deactivation rate constant. • The conversion versus time profile for aspartame synthesis by a batch process can be developed from this expression by integration. J.S. Parent
Effect of Thermolysin Instability on APM Production • The evolution of [L-Asp] and conversion with time for a batch process is shown below. • Depending on the relative rates of reaction and enzyme deactivation, the ultimate conversion can be strongly affected J.S. Parent
immobilized enzymes free (soluble) enzymes Effect of Immobilization on Operational Stability • Given that activity of enzymes is dictated by structure and conformation, the environmental change resulting from immobilization affects not only maximum activity, but the stability of the enzyme preparation. • The factors that inactivate enzymes are not systematically understood, and depend on the intrinsic nature of the enzyme, the method of immobilization, and the reaction conditions employed. • In general, immobilized enzyme preparations demonstrate better stability. • Note that the immobilized • preparation is often more • stable than the soluble • enzyme and displays a • period during which no • enzyme activity appears to • be lost. J.S. Parent
Classification of Immobilization Methods for Enzymes J.S. Parent
Immobilization by Entrapment • Gel entrapment places the enzyme within the interstitial • spaces of crosslinked, water-insoluble polymer gels. • Polyacrylamide gels: • Polysaccharides: The solubility of alginate and k-Carrageenan varies with the cation, allowing these soluble polymers to be crosslinked upon the addition of CaCl2 and KCl, respectively. • Variations of pore size result in enzyme leakage, even after washing. The effect of initiator used in polyacrylamide gels can be problematic. J.S. Parent
Immobilization by Entrapment • Microencapsulation encloses enzymes within spherical, • semi-permeable membranes of 1-100 mm diameter. • Urethane prepolymers, when mixed with an aqueous • enzyme solution crosslink via urea bonds to generate membranes of varying hydrophilicity. • Alternatively, photo- • crosslinkable resins • can be gelled by • UV-irradiation. • Advantage of Entrapment • Enzymes are immobilized without a chemical or structural modification. A very general technique. • Disadvantage of Entrapment • High molecular weight substrates have limited diffusivity, and cannot be treated with entrapped enzymes. J.S. Parent
Immobilization by Carrier Binding • Attachment of an enzyme to an insoluble carrier creates an active surface catalyst. Modes of surface attachment classify carrier methods into physical adsorption, ionic binding and covalent binding. • Physical Adsorption: Enzymes can be bound to carriers • by physical interaction such as hydrogen bonding and/or • van der Waal’s forces. • the enzyme structure is unmodified • carriers include chitosan, acrylamide polymers and silica-alumina • binding strength is usually weak and affected by temperature and the concentration of reactants. • Ionic Binding: Stronger enzyme-carrier binding is obtained with solid supports containing ion-exchange residues. • cellulose, glass-fibre paper, polystyrene sulfonate • pH and ionic strength effects can be significant J.S. Parent
Immobilization by Carrier Binding • Covalent attachment of soluble enzymes to an insoluble support is the most common immobilization technique. • Amino acid residues not involved in the active site can be used fix the enzyme to a solid carrier • Advantages: • 1. Minimal enzyme leaching from the support results • in stable productivity • 2. Surface placement permits enzyme contact with • large substrates • Disadvantages: • 1. Partial modification of residues that constitute the active site decreases activity • 2. Immobilization conditions can be difficult to optimize (often done • in the presence of a competitive inhibitor) J.S. Parent
Most Convenient Residues for Covalent Binding • Abundance(%)Reactions • 7.0 27 • 3.4 31 • 3.4 16 • 2.2 13 • 4.8 4 • 4.8 4 • 3.8 6 • 1.2 7 • Amino acid residues with polar and reactive functional groups are best for covalent binding, given that they are most often found on the surface of the enzyme. • Shown are the most convenient residues in descending order. The average percent composition of proteins (reactive residues only) is shown, along with the number of potential binding reactions in which the amino acids partake. J.S. Parent
Covalent Attachment Techniques • Cyanogen bromide activates supports with vicinal hydroxyl groups (polysaccharides, glass beads) to yield reactive imidocarbonate derivatives: • Diazonium derivatives of supports having aromatic amino groups are activated for enzyme immobilization: • Under the action of condensing agents (Woodward’s reagent K), carboxyl or amino groups of supports and amino acid residues can be condensed to yield peptide linkages. • Other methods include diazo coupling, alkylation, etc. J.S. Parent
Immobilization by Crosslinking • Bi- or multi-functional compounds serve as reagents for intermolecular crosslinking of enzymes, creating insoluble aggregates that are effective heterogeneous catalysts. • Reagents commonly have two identical functional groups • which react with specific amino acid residues. • Common reagents include glutaraldehyde, • and diisocyanates, • Involvement of the active site in crosslinking can lead to great reductions in activity, and the gelatinous nature of the product can complicate processing. J.S. Parent
Effects of Enzyme Immobilization on Activity J.S. Parent
Selecting an Immobilization Technique • It is well recognized that no one method can be regarded as the universal method for all applications or all enzymes. Consider, • widely different chemical characteristics of enzymes • different properties of substrates and products • range of potential processes employed J.S. Parent