Download

1 / 49
500 likes | 649 Vues
Chapter 9 The Mutability and Repair of DNA.

E N D
Chapter 9 The Mutability and Repair of DNA • As we all know that , the perpetuation of the genetic material from generation to generation depends on maintaining rates of mutation at low levels, or the high rates of mutation would destroy the species in the germ line and the individual in the soma.
At the same time ,if the genetic material were perpetuated with perfect fidelity the genetic variation needed to drive evolution would be lacking ,and new species would not arisen. • Therefore ,life and biodiversity depend in a happy balance between mutation and its repair , which is the main content of the chapter 9.
OUTLINE • Foreword • Replication errors and their repair • DNA damage • Repair of DNA damage • Something unknown • Summary
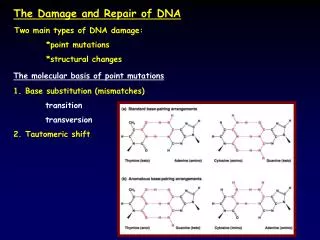
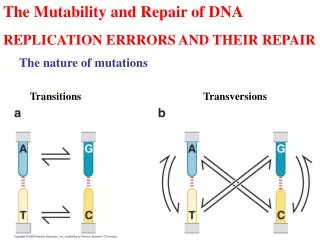
Foreword 1.Mutation (1)Definition Mutations include almost every conceivable change in DNA sequence. (2)type point mutation is the mutation that alter a single nucleotide . a . Transitions (pyrimidine to pyrimidine, purine to purine) b .Transversions (pyrimidine to purine, purine to pyrimidine
Insertions Deletions Gross rearrangement of chromosome “hotspots”: the sites on the chromosome where the mutations arise at high frequency. (eg. DNA microsatellite ,Mutation-prone sequence in human genome are repeats of simple di-, tri- or tetranucleotide sequences, these sequences (1) are important in human genetics and disease, (2) hard to be copied accurately and highly polymorphic in the population )
3.Two consequences of mutation unrepaired(1)Permanent changes to DNA which can alter the coding sequence of a gene or its regulatory sequence.(2)Some chemical alterations to DNA prevent its use as a template for replication and transcription.So repair of DNA is important to the organisms.OUTLINE
Replication errors and their repair As we have learned ,the proofreading mechanism ( the 3’→5’ exonuclease component) improves the fidelity of DNA replication, but it is not foolproof. So there are a total of 12 possible mismatches during the DNA replication (T:T,T:G,T:C, and so forth)
The misincorporated nucleotide needs to be detected and replaced, otherwise it will cause mutation.
Mismatch repair removes errors that escape proofreading Now let us discuss the process of repair in detail. Firstly,E. coli’s repair system 1.How to scan the mismatches and remove errors? 2.How to distinguish the mismatched strand and the parental strand?
MutS scans the DNA, recognizing the mismatch from the distortion they cause in the DNA backbone. MutS is a dimer of the mismatch repair protein. 1.How to scan the mismatches and remove errors?
MutS embraces the mismatch-containing DNA, inducing a pronounced kink in the DNA and a conformational change in MutS itself.
MutS-mismatch-containing DNA complex recruits MutL, MutL activates MutH, an enzyme causing an incision or nick on one strand near the site of the mismatch. MutL is a second protein component of the repair system. MutH is an enzyme causing an incision or nick on one strand
The helicase (UrvD) unwinds the DNA starting from the incision and moving in the direction of the site of the mismatch , and the exonucleases progressively digests the displaced single strand , extending to and beyond the site of the mismatched nucleotid. This action produces a single-stranded gap , which is then filled in by DNA polymerase Ⅲ and sealed with DNA ligase.
2.How to distinguish the mismatched strand and the parental strand? The answer is that E. coli tags the parental strand by transient hemimethylation as we now decribe.
The GATC sequence is widely distributed along the entire genome ,and all of these sites are methylated by the Dam methylase. So before the newly strand is methylated by the Dam methylase after the DNA replication ,the resulting daughter DNA duplexes will be hemimethylated , thus the newly strand is marked (it lacks a methyl group) and hence can be recognized as the strand for repair .
Different exonucleases are used to remove ssDNA between the nick created by MutH and the mismatch dependoing on whether MutH cuts the DNA in the 5’ or 3’ side of the misoncorporated nucleotide .
Secondly, the Eukaryotic cells’repair system • The Eukaryotic cells repair mismatches and do so using homologs to MutS (MSH) and MutL (MLH). • But they lack MutH and E. coli’s clever trick of using heminrthylation to tag the parental strand .
How does the mismatch repair system of the Eukaryotic cells know which of the two strands to correct ? As we see in chapter 8, takes place discontinuously wiyh the formation of Okazaki fragments that are joined to previously synthesized DNA by DNA ligase. Prior to the ligation step ,the Okazaki fragment is separated from previously synthesized DNA by a nick created, which can be though of as being equivalent to the nick created in E.coli by MuntH on the newly synthesized strand.OUTLINE
DNA undergoes damage spontaneously from hydrolysis and deamination. DNA damaged by alkylation , oxidation and radiation. Mutations are also caused by base analogs and intercalating agents. DNA damage
The most frequent and important kind of hydrolytic damage is deamination of the base cytosine, (just show in the picture on the left) Notice that , in contrast to the replication errors ,all of these hydrolytic reactions result in alterations to the DNA that are unnatural. 1.Hydrolytic damage
2.DNA damaged by alkylation , oxidation and radiation • Alkylation In alkylation ,methyl or ethyl groups are transferred to reactive sites on the baese and to phosphates in the DNA backcone.
Oxidation DNA is also subject to attack from reactive oxygen species .(eg. O2-, H2O2, and OH•) ■ radiation ultraviolet light ,which produce the photochemical fusion of two pyrimidines that occupy adjacent positions on the same polynucleotide chain. (eg. thymine dimer) These linked bases are incapable of base-pairing and cause the DNA polymerase to stop during replication. gamma radiation and X-ray, which cause double-strand breaks in the DNA ,which are different to repair.
3.caused by base analogs and intercalating agents • Base analogs Similar enough to the normal bases to be processed by cells and incorporated into DNA during replication. But they base pair differently, leading to mistake during replication.
Intercalating agents They are flat molecules containing several polycyclic rings that rings that bind to the equally flat purine or pyrimidine bases of DNA, just as the bases bind or stack with each other in the double helix. OUTLINE
Repair of DNA damage DNA repair system • Direct reversal of DNA damage • Excision repair system • Recombination (DSB) repairs • Translesion DNA synthesis
1. Direct reversal of DNA damage • A repair enzyme simply reverses (undoes) the damage • Now we will discuss two examples in detail to understand the direct reversal of DNA damage.
(1).Photoreactivation • The enzyme DNA photolyase captures energy from light and it to break the covalent bonds linking adjacent pyrimidines, so the damaged bases are mended directly.
(2).The removal of the methyl group The methyltransferase removes the methyl group from the methylated O6-methylguanine . The methyl group is transferred to the protein itself, inactivating the protein. (very costly)
2. Excision repair system • Two kinds of excision repair exist, one involving the removal of only the damaged nucleotide ,and the other ,the removal of a short stretch of single-strand DNA that contains the lesion.
(1).Base Excision repair • An enzyme called a glycosylase (lesion-specific) recognizes and removes the damaged base by hydrolyzing the glycosidic bond. • The resulting abasic sugar is removed from the DNA backbone. • After the damaged nucleotide has been entirely removed from the backbone, a repair DNA polymerase and DNA ligase restore an intact strand using the undamaged strand as a template.
What if a damaged base is not removed by base excision before DNA replication ? • There are some fail-safe systems to deal with this problem. • Then we will discuss two examples in detail.
oxoG mispair with A A dedicated glycosylase which recognizes the oxoG:A base pairs recognizes an A opposite an oxoG as a mutation and removed the undamaged but incorrect base.
T mispair with G • A glycosylase removes T from T:G mispairs. • The glycosylase system assumes that the T in the T:G mismatch arose from deamination of 5-methyl-cytosine and selectively removes the T so that it can be replaced with a C.
(2). Nucleotide Excision repair • The nucleotide Excision repair enzymes do not recognize any particular lesion, they work by recognizing distortions to the shape of the double helix. • Such distortions trigger a chain of events that lead to the removal of short single-stranded segment which is filled in DNA polymerase using the undamaged strand as a template .
The nucleotide Excision repair of E.coil (a)UvrA and UvrB scan DNA to identify a distortion
(b) UvrA leaves the complex ,and UvrB melts DNA locally around the distortion (c)UvrC forms a complex with UvrB and creates nicks to 5’ side of the lesion and to the 3’ side of the lesion . (d)DNA helicase UvrD releases the single stranded fragment from the duplex ,and DNA pol 1and ligase repair and seal the gap
Transcription-coupled repair nucleotide excision repair (NER) system is capable of rescuing RNA polymerase that has been arrested by the presence of lesions in the DNA template
3. Recombination (DSB) repairs • How do cells repair double-strand breaks in DNA in which both strands of the duplex are broken ? • 1.When the sister of the broken chromosome is present in the cell , the DSB-repair (double strand break system) pathway can operate. the DSB-repair retrieves sequence information from the sister chromosome. (Details are in chapter 10)
2.When a chromosome break early in the cell cycle, before a sister has been generated by DNA replication , a fail- safe system comes into play known as NHEJ (nonhomologous end joining) . NHEJ does not involve homologous recombination, instead, the two ends of broken DNA are directly joined to each other by misalignment between single strand protruding from the broken ends.
4. Translesion DNA synthesis • If cells cannot repair some lesions, there is a fail-safe mechanism that allows the replication machinery to bypass these sites of damage. This mechanism is known as translesion synthesis. • But because of its high error rate, translesion synthesis can be considered a system of last resort.
Translesion synthesis is catalyzed by a specialized class of DNA polymerases that synthesize DNA directly across the site of the damsge. OUTLINE
Something unknown • Although the repairs of damaged DNA have formed systems, there are still a lot of problems which have not be solved and need to be studied more deeply.
For example: • The mechanism by DNA glycosylase scan for damaged bases remains mysterious • In translesion synthesis 1.How does the translesion polymeras recognize a stalled replication fork ? 2.How does the translesion enzyme replace the normal replication polymerase in the DNA replication complex? 3.Once DNA synthesis is extended across the lesion ,how does tge normal replication polymerase switch back to and replace the translesion enzyme at the replication fork? OUTLINE
Summary • Now we have learned the whole knowledge on the mutation and repair of DNA ,so we would answer the questions as follows: 1.How is the DNA mended rapidly enough to prevent errors from becoming set in the genetic material as mutation? 2.How does the cell distinguish the parental strand from the daughter strand in repairing replication errors?
3.How does the cell restore the proper DNA sequence when the original sequence can no longer be read? 4.How does the cell deal with lesions that block replication?
The main content of this chapter: we have discussed errors that are generated during replication , lesions that arise from spontaneous damage to DNA , and the damage that is wrought by chemical agents and radiation. In my opinion , in each case we should consider what cause the alteration to the genetic material , how the alteration to the genetic material is detected and how it is properly repaired. Then if we understand these problems clearly, we must make it!
生科类.颜芳.200431060164 THE END! THANK YOU!