Download
1 / 17
170 likes | 192 Vues
This chapter discusses the reversible binding of proteins to ligands, with a focus on oxygen-binding proteins such as myoglobin and hemoglobin. It also explores the structure and function of the heme group in oxygen transport.

E N D
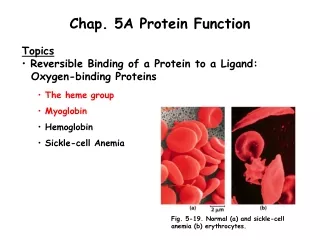
Chap. 5A Protein Function • Topics • Reversible Binding of a Protein to a Ligand: • Oxygen-binding Proteins • The heme group • Myoglobin • Hemoglobin • Sickle-cell Anemia Fig. 5-19. Normal (a) and sickle-cell anemia (b) erythrocytes.
Intro. to Ligand-binding Proteins Ligands are molecules that can be bound reversibly by a protein. Ligands can be any type of molecule, including another protein. Proteins that bind ligands do so at sequences called the binding site. The binding site is complementary in shape to the ligand that is bound. The degree of complementarity determines the binding specificity and strength. Most proteins undergo conformational changes on binding to a ligand. Changes can be small (i.e., “breathing”), or can be major. In a multisubunit protein, a conformational change in one subunit can affect the conformation of other subunits. Enzymes also bind to complementary molecules. However, in addition to binding, enzymes chemically transform these components. The binding site on an enzyme is referred to as the active, or catalytic site. The molecule acted on by the enzyme is referred to as the substrate.
Evolution of the Heme Group Molecular oxygen (O2) is only slightly soluble in aqueous solution. It cannot be carried to tissues in sufficient quantity if it is simply dissolved in blood plasma. Diffusion of O2 through the tissues is also ineffective over distances of greater than a few millimeters. For these reasons the evolution of large, multicellular animals depended on the evolution of proteins that could transport (e.g., hemoglobin) and store (e.g., myoglobin) oxygen. Still, something other than a protein per se is needed for O2 transport as none of the 20 standard amino acids can bind oxygen well. It is thought that this need led to the evolution of a bound prosthetic group know as heme which is functional in reversible O2 binding.
Structure and Properties of the Heme Group Heme consists of a complex organic ring structure known as protoporphyrin IX to which is bound a single iron atom in its ferrous (Fe2+) state (Fig. 5-1). The iron atom has six coordination bonds, four to nitrogen atoms that are part of the flat porphyrin ring (Fig. 5-1 d) and two that are perpendicular to the porphyrin (Fig. 5-1 b, c, & d). The coordinated nitrogen atoms help prevent conversion of the heme iron to the ferric (Fe3+) state which does not bind O2. Heme is found in many oxygen-transporting proteins as well as in proteins such as the cytochromes, which participate in electron-transfer reactions
Binding of the Heme Group to Myoglobin The heme group of myoglobin (and hemoglobin) is sequestered within a crevice, or pocket, in the protein (Refer to Fig. 4-16). The burying of the heme prevents a reaction that would occur with free heme in solution in which one O2 binds to two sandwiched heme groups and oxidizes iron to Fe3+. In myoglobin (and hemoglobin), only one of the perpendicular coordination positions of heme Fe2+ actually is unoccupied and available for O2 binding (Fig. 5-2). The other position is occupied by a His residue of the polypeptide chain that is commonly called the proximal histidine. When O2 binds to heme the electronic properties of the iron atom change and solutions containing the heme turn from a dark purple to a bright red color. Note that other small molecules such as carbon monoxide (CO), nitric oxide (NO), and cyanide ion (CN-) can replace O2 in the open coordination position. As discussed below, structural features of the heme binding pocket interfere with the binding of these other ligands and reduce their strength of interaction with heme iron.
The Globin Family The globins are a widespread family of proteins that are commonly found in eukaryotes of all classes and even in some bacteria. Most globins function in oxygen transport or storage, although some are involved in the sensing of gases such as O2, NO, and CO. At least four types of globins occur in humans and other mammals. These include the monomeric myoglobin, which facilitates oxygen diffusion in muscle tissue, and the tetrameric hemoglobin, which transports oxygen in the blood. A third monomeric globin, neuroglobin, is expressed in neurons and helps protect the brain from hypoxia (low oxygen) and ischemia (restricted blood supply). A fourth monomeric globin, cytoglobin, is found in number of tissues, but its function is unknown.
The Structure of Myoglobin Myoglobin (Mb) is a monomeric, globular protein (153 aas, Mr 16,700) that contains one heme prosthetic group. There are eight -helical segments in Mb and other globins, and these are labeled A through H consecutively in the sequence (Fig. 5-3). The bends in the molecule are designated AB, CD, EF, and so forth, based on the -helical segments they connect. Globins have an unusually high percentage of -helical structure (78% in Mb). In our discussion of Mb (and hemoglobin) structure and function, we will refer to several residues in the sequence. Note that these residues can be numbered based on their positions in the amino acid sequence, or by their locations in the sequence of a particular -helical segment. For example, the His residue that is coordinated to the heme group of Mb can be referred to as His93 or His F8.
Mathematical Description of Protein-ligand Binding (I) The reversible binding of a protein (P) to a ligand (L) can be described by the equilibrium expression P + L PL The reaction is characterized by an equilibrium constant, Ka, Ka = [PL]/[P][L] = ka/kd where ka and kd are rate constants for association and dissociation, respectively. Ka is the association constant that describes the equilibrium between the complex and the unbound components of the complex. The Ka provides a measure of the affinity of the ligand for the protein. Ka has units of M-1, and the higher the value of the Ka, the higher the affinity of the ligand for the protein. A rearrangement of the equilibrium equation shows that the ratio of bound to free protein is directly proportional to the concentration of the free ligand. Ka[L] = [PL]/[P]
Mathematical Description of Protein-ligand Binding (II) • The binding equilibrium can be described from the standpoint of the fraction (theta, ) of ligand binding sites in the protein that are occupied by ligand • = binding sites occupied/total binding sites = [PL]/([PL] + [P]) Substituting Ka[L][P] for [PL] and rearranging terms gives • = Ka[L][P]/(Ka[L][P] + [P]) = Ka[L]/(Ka[L] + 1) = [L]/([L] + 1/Ka) The value of Ka can be determined graphically from a plot of vs the concentration of the free ligand, [L] (Fig. 5-4a, next slide). Note that an equation of the form x = y/(y + z) describes a rectangular hyperbola, and therefore is a hyperbolic function of [L]. The fraction of ligand-binding sites occupied approaches saturation asymptotically as [L] increases. The [L] at which half of the available ligand-binding sites are occupied ( = 0.5) corresponds to 1/Ka.
Mathematical Description of Protein-ligand Binding (III) In the curve, the [L] at which half of the available ligand binding sites are occupied is equivalent to 1/Ka, or Kd (see next slide). The curve has a horizontal asymptote at = 1.0 and a vertical asymptote (not shown) at [L] = -1/Ka.
Mathematical Description of Protein-ligand Binding (IV) • It is more common (and intuitively simpler), to consider the dissociation constant, Kd, which is the reciprocal of Ka (Kd = 1/Ka) and is given in units of molar concentration (M). Kd is the equilibrium constant for the release of ligand from the complex. Expressions analogous to those derived above can be written using the Kd, namely • Kd = [P][L]/[PL] = kd/ka • and • = [L]/([L] + Kd) The latter equation plots the curve shown in Fig. 5-4a in the preceding slide. When [L] equals Kd, half of the ligand-binding sites are occupied. As [L] falls below Kd, progressively less of the protein has ligand bound to it. In order for 90% of the available ligand-binding sites to be occupied, [L] must be 9 times greater than the Kd. Finally, note that a lower value of the Kd corresponds to a higher affinity of the ligand for the protein (see Table 5-1, next slide).
Analysis of O2 Binding to Mb (I) A series of equations describing the binding of O2 to Mb can be derived from the equations used to describe the general binding of a ligand to a protein. First, the concentration of dissolved oxygen can be substituted for [L] into the equation relating to [L] and Kd, namely = [O2]/([O2] + Kd) As for any ligand, Kd equals the [O2] at which half of the available ligand-binding sites are occupied, or [O2]0.5. = [O2]/([O2] + [O2]0.5) In experiments using oxygen as a ligand, it is the partial pressure of oxygen (pO2) in the gas phase above the solution that is varied. pO2 is easier to measure than the concentration of O2 dissolved in solution. The concentration of a volatile substance in solution is always proportional to the partial pressure of the gas. So if we define the partial pressure of oxygen at [O2]0.5 as P50, substitution into the above equation gives = pO2/(pO2 + P50)
Analysis of O2 Binding to Mb (II) A binding curve for Mb that relates to pO2 is shown in Fig. 5-4b. The partial pressure of O2 in the air above the Mb solution is expressed in kilopascals (kPa). Oxygen binds tightly to Mb, with a P50 of only 0.26 kPa.
Protein Structure Affects Ligand Binding (I) The specificity with which heme binds its various ligands is altered when heme is a component of Mb (and hemoglobin). For example, CO binds to free heme molecules more than 20,000 times better than O2 (that is the Kd or P50 for CO binding to free heme is more than 20,000 times lower than that for O2). However, CO only binds about 200 times better than O2 when the heme is bound to Mb. This difference may be partly explained by steric hindrance (next slide). When O2 binds to free heme, the axis of the oxygen molecule is positioned at an angle to the Fe-O bond (Fig. 5-5 a). In contrast, when CO binds to free heme, the Fe, C, and O atoms lie in a straight line (Fig. 5-5 b). In both cases, the binding alignment reflects the geometry of the hybrid orbitals in each ligand. (Continued on the next slide).
Protein Structure Affects Ligand Binding (II) In Mb, His64 (His E7, the distal histidine) on the O2-binding side of the heme is too far away to coordinate with the heme iron, but it does interact with ligands bound to heme (Fig. 5-5 c). In fact, His E7 may help preclude the linear binding of CO, providing one explanation for the selectively diminished binding of CO to heme in Mb (and hemoglobin). The binding of O2 to heme in Mb depends on molecular motions, “breathing,” in the structure of the polypeptide chain. The heme group is buried so deeply in folded Mb that there is no direct path for O2 to the coordination site. In order for O2 to reach its binding site in heme, side-chains of Mb must move out of the way and produce transient cavities in the structure through which O2 can move.