ARO 2001
ARO 2001. A minimal model for the diversity of onset responses in the ventral cochlear nucleus. C.J. Sumner, R. Meddis, CNBH, Department of Psychology, University of Essex, Wivenhoe Park, Colchester, CO4 3SQ, U.K. contact: cjs@essex.ac.uk. abstract.


ARO 2001
E N D
Presentation Transcript
ARO 2001 A minimal model for the diversity of onset responses in the ventral cochlear nucleus C.J. Sumner, R. Meddis, CNBH, Department of Psychology, University of Essex, Wivenhoe Park, Colchester, CO4 3SQ, U.K. contact: cjs@essex.ac.uk
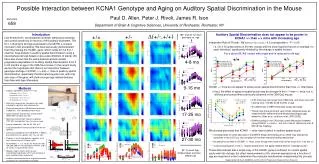
abstract There are neurons in the ventral cochlear nucleus that respond primarily at the onset of a pure tone stimulus. A number of mechanisms have been proposed as contributing to this characteristic: e.g. coincidence detection, inhibition, depolarisation block, and low-threshold potassium currents. We have investigated the minimum requirements of a model. We find that a simple point neuron having only a basic spiking mechanism, no special currents, and receiving only excitatory inputs from high-spontaneous rate auditory nerve (AN) fibers, is sufficient. The onset response arises from the convergence of a large number of small AN inputs, and the adaptation inherent in the auditory nerve response. Within the onset class, there is wide variation in observed responses. The following characteristics are reproduced by the model: 1. onset-I, onset-L and onset-C PSTH responses to BF-tones; 2. the ISIHs associated with these PSTH classes; 3. a very low deviation in first spike latency; 4. a diverse range of rate-intensity functions for BF-tones, including those with wide plateaus; 5. a range of wide and narrow receptive fields; 6. facilitation in two-tone frequency-threshold curves; 7. steep rate-intensity functions for broad-band noise stimuli. The models do not reproduce the observed range of rate-intensity functions for noise, which in vivo are as diverse as those for BF tones. The range of model responses arises from variations in membrane time constant, and the variation of innervation density with the characteristic frequency of AN inputs. The model therefore supports the coincidence detection hypothesis, and suggests that differences in excitatory innervation and integration time constant are sufficient to account for most of the variation in response characteristics. The failure of the model to reproduce the observed range of responses to broad-band noise, may reflect the lack of low-spontaneous rate fibers, or indicate a role for inhibition or extra currents.
I. Introduction INTRODUCTION • Onset neurons in the ventral cochlear nucleus (VCN) display a wide variety of responses, in terms of post-stimulus-time-histograms (PSTHs) 1,2, first order inter-spike interval histograms (ISIHs) 1,2, rate-intensity functions1,2, and other characteristics3. • Many mechanisms have been proposed as contributing to the response characteristics of these cells, particularly the onset response itself (e.g. inhibition4, coincidence detection1, depolarisation block5 , low threshold potassium currents5, stimulus dependant refractory effects9). • Onset cells display a wide range of receptive fields, some receiving inputs from AN fibres with a wide range of BFs. Probably because of this, they show a diverse range rate-intensity (RI) functions for BF-tones and broad-band noise. Their dynamic range is often much larger than AN fibres. Also thought to be linked to their wide receptive fields, is facilitation - a sub-threshold tone at BF can lower considerably the threshold of a tone presented away from the unit’s BF. • We are interested in establishing the minimum requirements of a model for the reproduction of these observed characteristics. Here we present a simple neural model architecture, with only a basic spiking mechanism. It uses no special currents or inactivation, and no inhibitory inputs, which might contribute to the generation of onset responses.
FINDINGS • The responses of a wide range of onset class neurons in the VCN can be modelled by changes to innervation alone. • The following characteristics have been modelled: • 1. PSTH BF-tone responses for the three classes of onset cell: pure-onset (onI), onset-chopper (onC) and onset-low (onL). • 2. The interval histograms associated with the PSTH classes. • 3. The variety of RI functions found in response to BF-tones. • 4. Saturating RI functions for broad-band noise. • 5. A variety of receptive fields. • 6. A variety of two-tone facilitation effects. • The model cannot reproduce the plateauing RI functions for broad-band noise. • The models employ no special currents or inhibition, and only receive input from high-spontaneous rate auditory-nerve fibres. These results therefore support the hypothesis that onset responses in the VCN can arise from coincidence detection alone. However, the rate-intensity functions for broad-band noise stimuli suggest that there may be a role for additional mechanisms in these cells. • The non-linear model of the cochlear supports medium- and low-spontaneous rate AN fibres. Using this we plan to investigate the effect of using inputs other than high-spontaneous rate fibres.
Auditory periphery: non-linear model of the guinea-pig cochlear 6 • AN transduction is by a revised version of the Meddis IHC (Sumner et. al. submitted). • The filter-bank has been tuned to reflect guinea-pig auditory nerve measurements (Evans 1972;Cooper and Yates 1993). • High-spontaneous rate fibres only (not a limitation of the filterbank). • Spike events are shaped by low-pass filtering to have a time course of about 1ms, and an asymmetric shape resembling somatic post-synaptic potentials. • The middle ear is modelled with a linear band-pass filter. • Mechanical filtering is modelled with a dual resonance non-linear (DRNL) filter-bank (Meddis et. al. 2001). Controlling the receptive field • The filter-bank is split into 5 contiguous groups. For each group, the CF range and number of fibres is specified separately. • Here, these variations to the receptive field are the principle way of varying the model. • The input current (I) size from each AN fibre is also varied in the model, but is constrained to be identical for all fibres.
Dendritic filtering • AN fibres synapsing on dendrites farther away (distal) are more attenuated and low pass filtered than those fibres synapsing closer to the soma (proximal). • This is simulated by separate low pass filtering of each group of shaped AN spikes. Filtering for each group can vary in attenuation, cut-off frequency and order. • Here only the ‘distal’ inputs are low pass-filtered (6th order, cut-off 100Hz). MacGregor neuron 7 • Simulates the basic properties of a spiking neuron. • Input is current injection of the sum of the dendritic filter outputs. • Fixed firing threshold (th0). • Absolute refractory period of 0.75ms. • Potassium hyper-polarization current with single exponential time constant (tGK = 0.2ms) • Single exponential membrane time constant (tm = 1ms). • Modified Hewitt and Meddis 8 (1993) chop-T stellate cell model. 0mV K+ hyper- polarization Membrane potential -50mV th0 -60mV f(tm & tGK) f(tm) + Iin -
III. PST and ISI responses to BF tones 1. Onset-chopper (OnC) • Three basic onset PSTH types have been found in the VCN: onI , onC and onL 1,2. They also have characteristic ISIHs. • The models successfully reproduce the different response type. This can be done with models that differ only by the patterns of AN innervation. • Only the distal AN inputs are low-pass filtered. Low-pass filtering confers a ‘dendritic delay’, which prevents inputs contributing to the onset part of the response. So dendritic inputs produce an on-L response. Data1 @ 50dB2 Data1 @ 20dB2 PSTH ISIH Spikes Model3 @ 20dB2 Model3 @ 50dB2 Spikes Time (ms) Time (ms) Modelparameters 1 all data is from Winter and Palmer1995 (120 repeats) 2 gains are with respect to unit threshold
2. Onset-low (OnL) 3. Onset-ideal (OnI) Data1 Data1 @ 60dB2 Data1 @ 30dB2 PSTH ISIH PSTH Spikes Spikes Model3 @ 30dB2 Model3 @ 50dB2 Model3 @ 60dB2 Spikes Spikes Time (ms) Time (ms) Time (ms) Modelparameters 3 model responses are made with 100 repeats
IV. Rate-Intensity functions 1. the data : RI functions for BF tones and broadband noise Steep for BBN • Onset cells have a display a diverse range of rate-intensity (RI) functions in response to BF tones and broad band noise (BBN). • RI functions for BF tones are normally sloping functions, but often have plateaus corresponding to a region of pure onset response (1 spike per stimulus presentation). • BBN RI functions can be similar to BF-tone responses, and can additionally show steep RI functions. • Here we compare the look at the variety of responses produced by the model, and how they depend on the adding of proximal and distal fibres to the on-I model. Sloping for BF-tone and BBN Sloping for BF-tone plateaus from Winter and Palmer (1995);Solid line: BF tone response; Dotted line: broad band noise response.
2. the model: RI functions for BF-tones • The model succeeds in reproducing the responses seen to pure tones at BF, using only HSR fibres. • AN inputs with BFs far from the neuron BF affect only high intensities. • AN inputs with BFs near to neuron BF affect low intensities. adding proximal HSR fibres (< 3/4 oct from neuron BF) adding distal HSR fibres (>3/4 oct from neuron BF) plateaus response of on-I unit 3. the model: RI functions for broadband noise adding distal HSR fibres (> 3/4 oct from neuron BF) • Responses of the model to broad-band noise all display steep saturating rate-intensity functions. • Effects are similar for proximal and distal inputs
1. the data : V. Receptive fields and facilitation • Onset neurons show a wide range of receptive fields, spanning up to 3 octaves. In the model these receptive fields arise from the variation in AN innervation. • Onset neurons often show facilitation: the threshold for two sub-threshold tones of different frequencies is lower than for one tone alone (Jiang et al. 1996). • Facilitation can be measured using two-tone frequency threshold curves (two-tone FTCs): FTCs for one tone are measured in the presence of a second tone, at BF, which does not on its own illicit a response. • In the model facilitation arises from the sub-threshold summation of AN inputs responding to the different tones. • Like the PSTHs, RI functions, and receptive fields, the nature of facilitation varies with the pattern of AN fibre inputs and their CFs. 2. the model :
Examples of guinea-pig two-tone FTCs (Jiang et. al.1996) Response area for a single tone: FTC Response area for second tone Sub-threshold BF tone Examples of model FTCs and two-tone FTCs Response area for a single tone: FTC Response area for second tone
references 1 Rhode WS, Smith PH, J Neurophysiol, 1986; 56, 261-286. 2 Winter IM, Palmer AR, J Neurophysiol, 1995; 73, 141-159. 3 Jiang D, Palmer AR, Winter IM, J Neurophysiol, 1996; 75, 380-395. 4 Paolini AG, Clark GM, J Neurophysiol, 2000; 81, 2347-2359. 5 Cai Y, Walsh EJ, McGee J, J Neurophysiol, 1997; 78, 872-883. 6 Meddis R, Hewitt M, J Acoust Soc Am, 1991; 89, 2866-2882. 7 MacGregor RJ, Neural and Brain Modelling, 1987; Academic, San Diego, CA. 8 Hewitt M, Meddis R, J Acoust Soc Am, 1992; 93, 3390-2279. 9 Kalluri S, Deglutte B, ARO 2000, # 639. acknowledgements Thanks to Ian Winter, and to L.P.O’Mard for the DSAM (Development System for Auditory Modelling) library upon which the models were implemented.