Download

1 / 136
1.4k likes | 1.66k Vues
Chapter 13: RNA Processing and Post-Transcriptional Gene Regulation. The discovery of split genes along with some other findings of recent years, shows that the genetic apparatus of the cell is more complex, more variable, and more dynamic than any of us had suspected.

E N D
Chapter 13: RNA Processing and Post-Transcriptional Gene Regulation
The discovery of split genes along with some other findings of recent years, shows that the genetic apparatus of the cell is more complex, more variable, and more dynamic than any of us had suspected. Pierre Chambon, Scientific American (1981) 244:60
Scientists who study RNA have been faced with more revolutionary and unexpected discoveries in the past several decades than in any other area of molecular biology. • A recurring theme in processing events that involve RNA cofactors is that the RNA is used to provide specificity by complementary base pairing.
1977: The discovery of split genes, first in adenovirus and then in a number of cellular genes. • Example: The ovalbumin gene was shown by R loop mapping to be split into eight sets of sequences.
1978: Walter Gilbert coined the term intron for the intervening sequences that split genes. • Until the early 1990s, the view was that introns are “junk” that is excised and degraded.
Now known that introns can be functional • May contain transcriptional regulatory elements. • May code for small nucleolar RNAs and microRNAs.
Modern definitions • Introns are sequences that remain physically separated after excision. • Exons are sequences that are ligated together after excision.
RNA splicing is the process by which introns are removed from a primary RNA transcript at precisely defined splice sites.
Intron-encoded small nucleolar RNAs and “inside-out” genes • snoRNAs mediate base modification of ribosomal RNA (rRNA) and possibly other RNAs. • By complementary base pairing with the rRNA, the snoRNAs recruit the modification enzymes to their target sites.
Box C/D snoRNAs guide 2′-O-methylation. • H/ACA snoRNAs direct pseudouridine formation.
In some “inside-out” genes, the introns code for function and the exons are degraded. • In 1994, U22 snoRNA and seven other snoRNAs were discovered within separate introns of the U22 host gene.
Five major classes of introns are distinguished by their structure and mechanisms of splicing • Autocatalytic group I introns • Autocatalytic group II introns • Archael introns • tRNA introns • Spliceosomal introns in nuclear pre-mRNA
Group I and group II self-splicing introns • Large catalytic RNAs distinguished by their structure and mechanisms of splicing. • Both types of introns have mobile members.
Group I introns require an external G cofactor for splicing • First characterized in Tetrahymena. • Widely distributed in the mitochondrial, chloroplast, and nuclear genomes of diverse eukaryotes. • In animals, group I introns have only been found so far in the mitochondrial genomes of one sea anemone and a coral.
Two-step splicing reaction, involving two transesterification reactions • Attack by an external guanine on the 5′ splice site, adding the G to the 5′ end of the intron and releasing the first exon. • The first exon attacks the 3′ splice site, ligating the two exons together and releasing the linear intron.
Although many group I introns can self-splice in vitro, many, if not all, require proteins in vivo to fold into the catalytically active structure. • Proteins required for splicing are either encoded by the introns themselves or by other genes of the host organism.
Group I intron structure • The typical secondary structure is approximately 10 base-paired helical elements organized into three stacked domains.
Group II introns require an internal bulged A for splicing • Less common than group I introns. • Found in the mitochondrial and chloroplast, genomes of certain protists, fungi, algae, and plants, and in bacterial genomes.
First transesterification reaction • Attack by the 2′-OH of an internal bulged A on the 5′ splice site. • Release of the first exon. • Formation of a lariat structure with a 2′→5′ phosphodiester bond.
Second transesterification reaction • The first exon attacks the 3′ splice site. • The two exons are ligated together. • The lariat intron is released, debranched, and degraded.
Although many group II introns can self-splice in vitro, many, if not all, require proteins in vivo to fold into the catalytically active structure. • Proteins required for splicing are either encoded by the introns themselves or by other genes of the host organism.
Group II intron structure • Conserved tertiary structure. • Six major domains radiating from a central wheel.
Mobile group I and group II introns • Spread efficiently into a homologous position in an allele that lacks the intron. • Movement is mediated by highly-specific homing endonucleases that are typically encoded by the self-splicing intron. • Group II introns also encode an open reading frame with homology to reverse transcriptase.
Group I introns typically move by a process termed homing. • Group II introns typically move by a process termed retrohoming.
Archael introns are spliced by an endoribonuclease • Archaea carry introns in their tRNA, rRNA, and mRNA that are spliced by an archael-specific mechanism. • Cut-and-rejoin mechanism that requires ATP, an endoribonuclease, and a ligase. • Bulge-helix-bulge motif at the exon-intron junction.
Some nuclear tRNA genes contain an intron • Presence or absence of an intron defines two classes of eukaryotic nuclear tRNA genes. • In humans, only tRNATyr and tRNALeu contain introns.
Nuclear tRNA splicing involves two main reactions: • Cleavage: The intron-containing pre-tRNAs are cleaved by an endoribonuclease at the 5′ and 3′ boundaries of the intron. • Joining: The paired tRNA halves are joined by ligase.
In plants and fungi, the 2′ phosphate and 3′-OH at the splice junction are resolved by phosphotransferase.
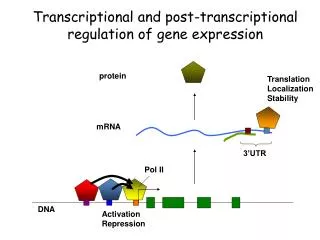
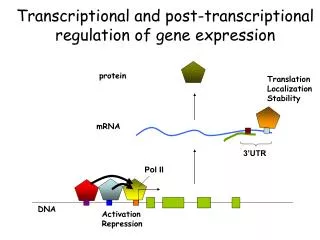
Eukaryotic mRNA is covalently processed in three ways prior to export from the nucleus: • Transcripts are capped at their 5′ end with a methylated guanosine nucleotide. • Introns are removed by splicing. • 3′ ends are cleaved and extended with a poly(A) tail.
Cotranscriptional processing of nuclear pre-mRNA • The C-terminal domain (CTD) of RNA polymerase II functions as a “landing pad” for RNA processing factors. • Deletion of the RNA polymerase II CTD inhibits capping, splicing, and poly(A) cleavage.
Experiments have shown that different regions of the CTD serve distinct functions in pre-mRNA processing. • The C-terminal heptapeptide repeats support capping, splicing, and 3′ processing. • The amino-terminal repeats only support capping.
Addition of the 5′-7-methylguanosine cap • The cap protects mRNA from degradation and enhances the efficiency of splicing, nuclear export, and translation. • A distinguishing chemical feature of the cap is the 5′→5′ linkage of 7-methylguanosine to the initial nucleotide of the mRNA. • Often abbreviated as m7GpppN to reflect this linkage.
The cap is added in three main steps: • An RNA triphosphatase removes the terminal phosphate from the pre-mRNA. • Guanylyltransferase adds the capping GMP from GTP. • Methyltransferases methylate the N-7 of the capping guanosine and the 2′-O-methyl group of the penultimate nucleotide.
Termination and polyadenylation • No consensus termination sequence has been identified. • Most metazoan mRNA 3′ ends are produced by cleavage of the pre-mRNA at the polyadenylation site between conserved AAUAAA and G/U-rich sequence elements. • These regions are recognized by cleavage and polyadenylation specificity factor (CPSF) and cleavage stimulation factor (CstF).
Cleavage requires two additional complexes, mammalian cleavage factor I and II, CFIm, CFIIm. • The mRNA is cleaved while it is still being synthesized. • Cleavage also occurs at a cotranscriptional cleavage site (CoTC) which, at least in the case of the -globin mRNA, is self-cleaving.
The remaining transcript still associated with RNA polymerase II is attacked by the Xrn2 exonuclease that chases after the polymerase. • When the exonuclease catches up, transcription is terminated.
The catalytic CoTC core of the -globin pre-mRNA folds into a defined secondary structure. • The minimal catalytic core was shown to self-cleave in a time course conducted under protein-free conditions.
After cleavage and release of the mRNA, the transcript is polyadenylated at the 3′ end. • Most eukaryotic mRNAs have a chain of A residues about 100 to 250 nt long. • Histone mRNAs have a conserved stem-loop structure instead of a poly(A) tail.
The poly(A) tail is added by poly(A) polymerase. • Enhances mRNA stability and translation efficiency.
The poly(A) tails are coated with sequence-specific poly(A)-binding proteins. • In the nucleus, PABPN1 increases the processivity of poly(A) polymerase. • In the cytoplasm, PABPC functions in the initiation of translation and the regulation of mRNA decay.
Oculopharyngeal muscular dystrophy: trinucleotide repeat expansion in a poly(A)-binding protein gene • Muscular dystrophy that begins in the eyes and throat. • Prevalence is highest in French-Canadians who are descended from French immigrants, a man and wife, who emigrated to Canada in 1634.
Trinucleotide repeat expansion in exon 1 of the PABPN1 gene. • Normal protein expanded from 10 alanines in a row to 12-17 alanines. • Leads to intranuclear protein aggregation.
Splicing • Basic mechanisms is the same as self-splicing group II introns. • Except, the 3-D structure required for splicing is generated by the spliceosome.
Components of the spliceosome • Five uracil-rich small nuclear RNA (snRNA)-protein complexes termed snRNPs. U1, U2, U4, U5, U6 What happened to U3? • >200 proteins.
The structure of snRNPs • snRNAs are associated with Sm or Sm-like common core proteins and particle-specific proteins. • Sm proteins are named after the patient whose autoimmune serum was first used to detect them. • Sm proteins form a doughnut-shaped complex.
Spinal muscular atrophy: defects in snRNP biogenesis • Fatal autosomal recessive diseases. • Degeneration of motor neurons in the anterior horn of the spinal cord. • Mutations in the Survival of Motor Neurons 1 (SMN) gene. • Defects in snRNP biogenesis.

![RNA interference (RNAi) [aka post-transcriptional gene silencing (PTGS)]](https://cdn4.slideserve.com/584681/rna-interference-rnai-aka-post-transcriptional-dt.jpg)




![[VI]. Post-Transcriptional Processing and Post-Transcriptional Control of Gene Expression](https://cdn1.slideserve.com/3214110/vi-post-transcriptional-processing-and-post-transcriptional-control-of-gene-expression-dt.jpg)