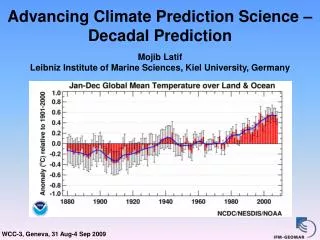
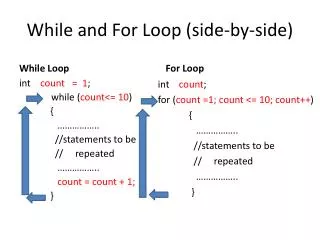
Download

1 / 0
0 likes | 238 Vues
موضوعات سمينار : Side chain dihedral roramer libraries Polarizable force fields Plop software Loop prediction Pk a prediction by simulation SMD simulation REMD simulation Δ G binding prediction by simulation Salvation model in simulation Inverse-docking

E N D