Download

1 / 31
350 likes | 746 Vues
Plasma Membrane, Lecture Outline 1. Function 2. Structure A. Phospholipid (PL) bilayer: bilayer organization; phospholipid composition; glycolipids and cholesterol; molecular structures; lipid rafts; B. Membrane proteins (MP): peripheral, integral, membrane anchored

E N D
Plasma Membrane, Lecture Outline • 1. Function • 2. Structure • A. Phospholipid (PL) bilayer: bilayer organization; phospholipid composition; glycolipids and cholesterol; molecular structures; lipid rafts; B. Membrane proteins (MP): peripheral, integral, membrane anchored • 3. Mechanisms of Transport across PM • A. Small molecules: Passive vs. facilitated diffusion vs. active transport • B. Macromolecules: Endocytosis • 1. Clathrin-mediated • 2. Clathrin-independent: caveolar uptake • 3. Ubiquitin-mediated 4. Machinery involved in Vesicular Transport and Fusion • A. Steps in vesicular targeting. • B. Cellular machinery involved in vesicular transport. • C. Mechanism of vesicular fusion. • 5. Examples from Pathobiology A.HIV-1 Nef as an adaptor in receptor mediated endocytosis • B. Viruses usurp cellular endocytosis machinery for budding • Pathogens that enter cells via caveolae avoid lysosmal fusion J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane (PM): 1. Functions: Defines the boundary of the cell and isolates the cell. Acts as a selective barrier - maintains composition of cytoplasm, which is very different from extracellular space. Mediates the interaction of the cell with its environment. Traversed by pathogens for access to the cell interior. Mammalian Cell Intracellular and Extracellular Ion Concentration J. R. Lingappa, Pabio 552, Lecture 6
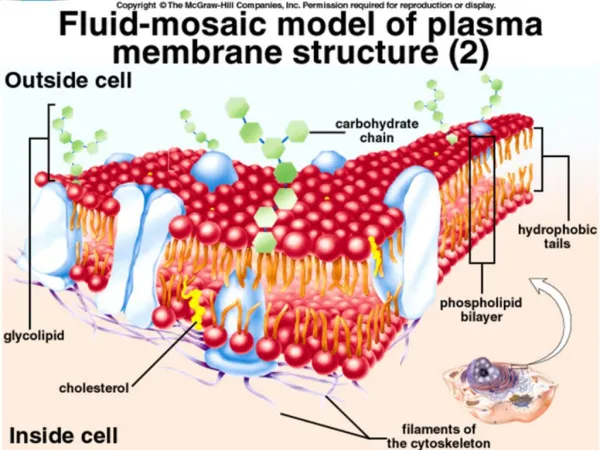
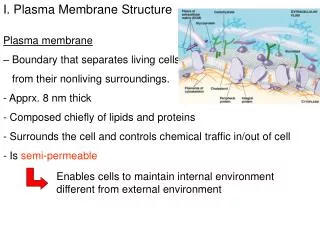
The Plasma Membrane (PM): 2. Structure: lipid bilayer ~ 5 nm thick A. Phospholipid (PL) bilayer - impermeable to water soluble molecules. 1. Importance of lipid bilayer organization. a. Hydrophobic fatty acid tails on inside b. Hydrophilic fatty acid heads on outside c. Viscous fluid: allows PLs and proteins to diffuse laterally within PM d. Caveats to fluid mosaic model: Rafts inhibit lateral mobility Flippase enzymes catalyze flipping to other half of bilayer J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane (PM): 2. Structure: lipid bilayer ~ 5 nm thick A. Phospholipid (PL) bilayer - impermeable to water soluble molecules. 1. The behavior of lipids: micelles vs. vesicles. J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane (PM): 2. Structure, cont.: A. Phospholid Bilayer.: 2. Phospholipid (PL) composition, mammalian cells: 4 major PL (~50% of PM lipid); 1 minor PL: • a. phosphatidylcholine (PC) & sphingomyelin (SM) - mainly in outer leaflet. • b. phosphtidylethanolamine (PE) & phosphatidylserine (PS) - inner leaflet • c. phosphatidylinositol (PI) - minor component in cytosolic leaflet but important for signaling. • d. PS and PI - negatively charged, giving net negative charge to cytosolic face. (note: E. coli = predominantly PE, no PC, PS, PI, SM, or cholesterol.) 3. Glycolipids: ~2 % of PM lipid; exclusively in out leaflet (non-cytosolic) 4. Cholesterol: rigid with polar hydroxyl group facing out. 5 % of PM lipid maintains membrane rigidity at high temps maintains membrane fluidity at low temps not present in bacteria Cholesterol with phospholipids Cholesterol J. R. Lingappa, Pabio 552, Lecture 6
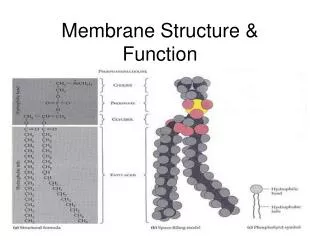
The Plasma Membrane (PM): 2. Structure, cont. A. Phospholipid Bilayer: 4. Molecular Structures: all are amphipathic a. PL = glycerol attached to 2 FA phosphate and different side groups (PE, PS, PC) b. SM = serine attached to 2FA phosphate and choline side group c. PI = minor phospholipid critical for signaling; inositol ring can be phosphorylated d. Cholesterol = complex hydrocarbon ring structure Phospholipids J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane (PM): 2. Structure, cont. A. Phospholipid Bilayer: 4. Phosphoinositol - has a ring structure that can be phosphorylated; also cleavage of this ring results in formation of 2 new structures that are both active in signaling (DAG and IP3). Phospholipase cleavage Signaling by PI in PM: J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane (PM): 2. Structure, cont. A. Phospholipid Bilayer: 4. Molecular Structures: all are amphipathic e. Glycolipids = lipid with sugar molecules attached Glycolipids J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane (PM): 2. Structure, cont.: A. Phospholipid Bilayer: 5. "Lipid rafts": dynamic regions of the plasma membrane enriched in cholesterol, sphingomyelin, glycolipids, GPI-anchored proteins and some membrane proteins. Important for signaling. Important as sites for entry and egress of viruses. Markers for clathrin-mediated endocytosis are not present in rafts. Insoluble in cold detergent; dispersed by cholesterol depletion (methyl-b-cyclodextrin). J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 2. Structure: B. Membrane associated proteins (MP): a. PM consists of 50% protein; 50% lipid. b. MP mediate selective traffic of molecules into and out of cell. c. Peripheral MP: dissociate from PM with high pH or high salt (carbonate extraction, pH 10). hydrophilic, assoc.via prot.-prot. interactions. d. Integral MP: released from PM only by solubilizing membranes with detergents. many are transmembrane proteins that span the bilayer. Membrane associated proteins (MP): J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 2. Structure: C. Use of Detergents: a. Non-ionic detergents solubilize membranes and membrane proteins without denaturing proteins b. Ionic detergents solubilize membranes and denature proteins J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 2. Structure: C. Use of Detergents: Vesicle Reconstitution J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: Mechanisms for Transport across the PM: A. Small molecule transport: 1. Passive diffusion: no MP involved. small hydrophobic molecules. 2. Facilitated diffusion: mediated by MP, but not energy-dependent. e.g.: glucose and amino aicds (via carrier proteins) and charged ions such as H+, Cl-, Na+, Ca+ (via channels). 3. Active transport: transport against concentration gradient, driven by ATP hydrolysis. e.g.: Na+-K+ pump, Ca+ pump, ABC transporters. J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 3. Mechanisms for Macromolecule Transport across the PM: B. Endocytosis: 1. Clathrin-mediated: a. Receptors mediate binding to ligands (lipids, ligands, sol. proteins, viruses). b. Selection of receptor or receptor-ligand for transport: "sorting signal" in receptor tail interacts w/ cytosolic adaptor to form "assembly particles" (AP) that interact w/ clathrin. c. Vesicles form by clathrin polymerization & using reg. proteins (dynamin, ARFs). d. Vesicle targets to endosome. e. Exposure to acidic pH in early endosome in some cases dissociates ligand from receptor; in other cases no dissociation. f. Cargo, or receptor plus cargo: sent to lysosome for degradation. g. Dissociated receptor recycled to plasma membrane. J. R. Lingappa, Pabio 552, Lecture 6
Plasma Membrane: 3. Transport Across PM: B. Endocytosis, cont.: 1. Clathrin-mediated: Diagrams showing Clathrin assembly/ disassembly, and dynamin J. R. Lingappa, Pabio 552, Lecture 6
Plasma Membrane: 3. Transport Across PM: B. Endocytosis: 1. Clathrin-Mediated: Four types of endocytic sorting signals on cytoplasmic domain of membrane proteins that direct endocytosed proteins into clathrin-coated pits: a. tyrosine based signals, i.e. YXXf (f = large hydrophobic aa); adapter= AP2 b. dileucine (LL) -containing signals; adapter = AP2 c. phosphorylated serine rich domain at the C-terminus d. motifs that recruit mono-ubiquitination machinery; adaptors = Eps15/15R, epsins and Hrs. J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 3. Mechanisms for Transport across the PM, cont.: B. Endocytosis, cont.: 2. Caveolar uptake Caveolae: flask-shaped or flat, non-coated membrane invaginations, 50 - 100 nm Like lipid rafts: contain cholesterol, glycoshpingolipids, GPI-anch. proteins, receptors Unlike lipid rafts: contain caveolin-1: 178aa, TM protein; interacts w/signaling molecules Lipid rafts are the precursors for caveolae formation Centers for signalling activity as well as endocytosis Exclude receptors involved in clathrin-dependent uptake Cholesterol depletion perturbs rafts & caveolar uptake (not clathrin) Pinching off and delivery into caveosomes which are are much more stable than endosomes; these deliver cargo to ER, Golgi Site of entry for nutrients, hormones, chemokines; also selected viruses, bacteria, parasites, and bacterial toxins. Entry via caveolae allows pathogen to evade fusion with lysosomes and degradation. Left: EM of caveolae. Right: SV40 enters via caveolae and traffics to the ER. GPI -anchored proteins enter via caveolae & traffic to theGolgi Pfeffer, Nat. Cell Biol. 3:E108 (2001) J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 3. Mechanisms for transport of macromolecules, cont. B. Endocytosis, cont.: 3. Ubiquitin-mediated endoctyosis (UME): Ubiquitin: 76 aa protein that gets conjugated to substrate proteins Poly-ubiquitination targets proteins for degradation by proteasome. Mono-ubiquitination acts as a signal for endocytosis of proteins at the cell-surface. Cell surface residence for a specific time triggers internalization (ubiq-indep). Ubiquitinated receptors are internalized into endosomes, multivesicular bodies (MVB = late endosome), and the lysosome. Non-ubiquitinated receptors are recycled to the plasma membrane via recycling endosomes. Mono-ubiquitinated internalized proteins interact with endocytic adapter complexes through surface patches surrounding critical residues within ubiquitin. Adaptor proteins such as epsins have ubiquitin-interacting motifs (UIM) that reqcognize mono-ubiquitinylated proteins and interact with clathrin adaptor proteins. Sorting of Ub-substrates into endosomes, MVB, and lysosomes requires: interaction with ESCRT complexes containing Vps proteins (yeast) mammalian equivalents include Tsg101 and Hrs De-ubiquitinating enzymes remove Ub for recycling and re-use. J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane:3. Mechanisms for transport of macromolecules, cont. B. Endocytosis, cont.: 3. Ubiquitin-mediated endoctyosis (UME): J. R. Lingappa, Pabio 552, Lecture 6
Plasma Membrane: 4. Vesicle Transport and Fusion A. Steps in Vesicular Targeting: 1. Transport vesicle with v-SNARE is tethered to target mb by a Rab GTPase. 2. If v-SNARE on vesicle and t-SNARE on target match, then loosely tethered vesicle becomes tightly "docked". J. R. Lingappa, Pabio 552, Lecture 6
Plasma Membrane 4.Vesicular Transport & Fusion, cont.: A. Steps in Vesicular Targeting: 3. Fusion is facilitated by SNAREs. 4. The trans-SNARE complex (now cis-SNARE) is then disrupted by the action of NSF and SNAP, which are recruited to the complex after formation of the SNARE complex, making the SNAREs available to form new complexes. 5. Recycling of the v-SNARE back to the donor compartment. 6. Note that requirement for disassembly of SNARE complexes prevents indiscriminate fusion between membranes by introducing a regulatory step. J. R. Lingappa, Pabio 552, Lecture 6
Plasma Membrane 4. Vesicle Targeting and Fusion, cont.: B. Machinery Involved: Rab-GTPases - small GTP binding proteins on vesicles. Related to the oncogene product Ras. Act as tethering factors that mediate initial interaction between membranes. Bind to Rab effectors on target membrane. Over 30 diferent Rab proteins specific to different membranes. Another protein (guanine-nuc. exchange factor) catalyzes exchange of GDP bound to cytosolic Rab for GTP, which allows Rab to bind to the transport vesicle. NSF - (N-ethylmaleimide sensitive factor) a tetramer of identical subunits that binds and hydrolyzes ATP. Required for disassembly of SNARE complex. SNAPs - (soluble NSF attachment protein). Act as a cofactor mediating NSF attachment to SNAREs. SNAP-NSF Receptors (SNAREs) - a family of cognate membrane proteins. Vesicular (v)-SNAREs on vesicles form complexes with target (t)-SNAREs on target membranes, either on the same membrane (cis) or different membranes (trans). SNAREs alone can cause fusion of membranes, although most likely in cells they act as direct catalysts of fusion along with other regulatory and triggering proteins. J. R. Lingappa, Pabio 552, Lecture 6
Plasma Membrane 4. Vesicle Targeting and Fusion: C. Fusion Mechanism: 1. Docking and fusion are separate steps. 2. Fusion involves displacement of water and lipids flowing from one bilayer to the other. 3. SNARE complexes may squeeze out water molecules and pull lipid bilayers together to form fusion intermediates. 4. SNAREs are the minimal machinery required for membrane fusion (how do you think these experiments were done?). 5. In vivo, other regulatory events, like calcium influx, may also be involved in triggering fusion. J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: • Examples from Pathobiology: J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 5. Examples from Pathobiology: B. Viruses usurp cellular endocytosis machinery for budding: HIV-1 and Ebola use cellular proteins (Tsg101 and Vps4) involved in endocytic sorting of ubiquitinated proteins to facilitate budding of progeny virus from the cell. Note that this process is topologically identical to the budding events that occur when endosomes are converted into multivesicular bodies (MVB). In fact, Tsg101 and Vps4 are both used in uninfected cells for budding into the MVB. Machinery for Ubiquitin-Mediated Endocytosis used by HIV-1 and Ebola: Mark Marsh & Markus Thali Nature Medicine 9, 1262 - 1263 (2003) From Strous and Gent, FEBS lett. 529: 102 (2002) J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 5. Examples from Pathobiology: B. Viruses usurp cellular endocytosis machinery for budding: Virus can be delivered as a packet onto another cells when the MVB moves to the PM (in dendritic cells and macrophages). J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 5. Examples from Pathobiology: B. Viruses usurp cellular endocytosis machinery for budding: This form of virus delivery could lead to immunologically protected sites of virus delivery. J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 5. Examples from Pathobiology: C. Pathogens that enter cells via caveolae or lipid rafts can target to various intracellular compartments and avoid lysosomal fusion. • Includes viruses, bacteria, mycobacteria, and parasites. From Duncan et al. Cellular Microbiology 4: 783 (2002) J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 5. Examples from Pathobiology: D. Influenza Virus Fusion: Virus binds to PM and is internalized by endocytosis Low endosomal pH induces conformation chage in HA leading to fusion of viral membrane with endosomal membrane, allowing virus to enter cell. HA protein of influenza: trimeric, integral membrane protein Monomer is HA0, which is cleaved post-translationally to produce HA1 & HA2 HA2 subunit in viral mb, HA1 largely distal Last 20-25 amino acids of HA2 are the fusion peptide (12aa, mostly hydrophobic) Upon cleavage of HA0: fusion peptide folds into pocket in stem Low pH causes conformation change in HA that exposes fusion peptide Fusion peptide then interacts with endosomal membrane and brings membranes together allowing fusion to occur J. R. Lingappa, Pabio 552, Lecture 2-30 Entire HA molecule HA2 subunits alone: conformational change exposes fusion peptide and membranes are brought together From Skehel and Wiley, Annu Rev Biochem. 2000;69:531-69. J. R. Lingappa, Pabio 552, Lecture 6
The Plasma Membrane: 5. Examples from Pathobiology: D. Influenza Virus Fusion, cont. J. R. Lingappa, Pabio 552, Lecture 2-31 J. R. Lingappa, Pabio 552, Lecture 6
Recommended Reviews on Pathogens and the Plasma Membrane: Marsh, M. and A. Helenius.Virus entry: open sesame.Cell. Feb 24;124(4):729-40. Review (2006). Sieczkarski, S. and G. Whittaker. Dissecting virus entry via endocytosis. J. Gen. Virology 83: 1535 (2002). van Deurs, B. et al. Caveolae: anchored, multifunctional platforms in the lipid ocean. Trends Cell Biol. 13: 92 (2003) Duncan, et al. Microbial entry through caveolae: variations on a theme. Cellular Microbiology 4: 783-91 (2002). Pelkmans, L. and A. Helenius. Endocytosis via caveolae. Traffic 3:311 (2002)). Bromsel, M. and A. Alfsen. Entry of viruses through the epithelial barrier: pathogenic trickery. Nat. Rev. Mol. Cell Biol. 4:57 - 68 (2003). J. R. Lingappa, Pabio 552, Lecture 6