lupus and complement deficiency : insights from knockout mice
300 likes | 740 Vues
Disclosures: No Relevant Financial Relationships with Commercial Interests. Complement and SLE - objectives. To explore the links between complement and break of B cell toleranceTo review the links between complement and clearance of dying cellsTo analyse the role of complement in tissue damageTo consider the background strain effects in gene-targeted mice.

lupus and complement deficiency : insights from knockout mice
E N D
Presentation Transcript
1. Marina Botto
2. Disclosures:
No Relevant Financial Relationships with Commercial Interests
3. Complement and SLE - objectives
To explore the links between complement and break of B cell tolerance
To review the links between complement and clearance of dying cells
To analyse the role of complement in tissue damage
To consider the background strain effects in gene-targeted mice
4. Complement ACTIVATION
5. Complement and SLE Active SLE is associated with hypo-complementaemia (reduced levels of C1q, C2, C4, CH50 and C3)
Complement levels increase following treatment
Tissue biopsies show deposits of C3 and C4
Complement activation can cause tissue damage
6. Pathogenesis of inflammation in SLE The traditional view
Autoantibodies form immune complexes with autoantigens
These fix complement
Complement causes tissue injury
but
The clinical evidence doesn�t quite fit!
7. Complement and SLE Clues from clinical observations
8. Complement deficiency and SLE
9. Complement - physiology Interface between innate & adaptive immunity
augmentation of antibody responses
C3 and C4 fragments bound to antigens and immune complexes
C3 receptors on B cells and APCs
enhancement of immunological memory
C3 and C4 fragments bound to antigens and immune complexes
C3 receptors on follicular dendritic cells
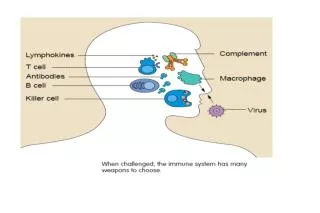
10. Importance of complement in immune response
11. The paradox complement deficiency is associated with reduced antibody responses and defective humoral immunity
complement deficiency is associated with SLE in which
increased antibody responses to self antigens
tissue injury mediated by immune complexes
12. Complement FUNCTION
13. Complement and SLE pathogenesis � 3 steps Complement and pathological survival of autoreactive lymphocytes
2) Complement and failure of clearing
dying cells
3) Complement and tissue damage
14. Does C1q have a direct role in the maintenance of B cell tolerance ?
15. C1q-deficiency and self tolerance C1q deficient mice bred with hen egg lysozyme (HEL)-transgenic mice expressing:
a) Ig H and L chains with high affinity for HEL
b) sHEL (10-30ng): sufficient to trigger anergy but not deletion of transgenic B cells
17. Serum anti-HEL antibody levels
18. C1q-deficiency and self tolerance 1) normal �self-tolerance� to sHEL as autoantigen in C1q deficiency
2) C1q deficient mice crossed with anti-ssDNA
knock-in transgenic mice (3H9R/Vk8R):
regulation of the single specificity anti-ssDNA
Anergy induction in both these models was not modified by C1q deficiency
19. SLE pathogenesis � 3 steps Complement and pathological survival of autoreactive lymphocytes
2) Complement and failure of clearing
dying cells
3) Complement and tissue damage
21. Does complement contribute to the clearance of dying cells?
22. In vivo binding of C1q to apoptotic keratinocytes
23. C1q deficient mouseGlomerular apoptotic bodies
26. How does C1q contribute to the clearance of apoptotic cells? C1q binds directly to apoptotic cells. C1q bound to apoptotic cells:
a) can activate the classical pathway with subsequent C4b and C3b/bi deposition
b) is recognised by calreticulin on phagocytes and this in conjunction with CD91 stimulates the uptake of apoptotic cells by pinocytosis
C1q binding to apoptotic cells is mediated by IgM antibodies and/or pentraxin and this causes activation of the classical pathway
28. Does the impaired clearance of complement deficiency have an effect on B cells?
29. C1q and B cell tolerance to intracellular antigens Tg mice expressing an intracellular membrane-bound form of HEL (mHEL-KK) that is sequestered in the ER of all MHC I expressing cells
mHEL bound on the cell surface induces tolerance by deletion/editing; sHEL induces tolerance by anergy
Intracellular mHEL-KK positively selects B1 cells and high titres of IgM autoantibodies. Intracellular self-antigens are immunogenic and can stimulate the conventional B cells that escape negative selection
C1q �deficient mice bred with Tg mice expressing mHEL-KK
30. Increased positive selection of IgM autoAb and peritoneal B1 cells in C1q-/- mice
31. Increased numbers of conventional B cells specific for mHEL-KK in C1q-/- mice
32. IgM autoantibody recognise intracellular antigens expressed on apoptotic cells
33. Phagocytosis of non-Tg and mHEL-KK apoptotic thymocytes
34. C1q and B cell tolerance to intracellular antigens - conclusions C1q inhibits the positive selection of B1b B cells and IgM autoantibodies by an intracellular self antigens
C1q contributes to the clearance of dying cells that may explain the increased positive selection of auto-reactive B1 cells by intracellular antigens
35. C1q, clearance of dying cells and SLE: hypothetical model
36. SLE pathogenesis � 3 steps Complement and pathological survival of autoreactive lymphocytes
2) Complement and failure of clearing
dying cells
3) Complement and tissue damage
37. What is the role of complement in glomerulonephritis?
38. C1q deficiency, alternative pathway deficiency and nephritis
39. Spontaneous glomerulonephritis and complement deficiency
40. C1q-deficient mice -conclusions C1q-deficient mice developed a lupus-like disease and had a defect in the clearance of dying cells that may explain the association between complement deficiency and SLE
C1q inhibits the positive selection of B1 B cells and IgM autoantibodies by an intracellular self antigens
41.
43. Do lupus-prone strains of mice have an impaired clearance of apoptotic cells?
46. Is a defective clearance of dying cells sufficient to induce SLE ?
49. Background strain effects on autoimmunity in knockout models of SLE Our lab is working on SLE (Systemic Lupus Erythematosus, or Lupus as it is commonly known). This is a multi-system autoimmune disease with a complex genetic basis. Murine models of SLE have proved useful in attempting to identify the genes involved.
Three major mouse models of the disease have been described: NZB/NZW hybrid, MRL lpr and BXSB.
Targeted disruptions of genes of immune system proteins have also been used (gene knockouts) and these seem to have a surprisingly high frequency of autoimmune phenotype associated with them.
Our lab has generated several knockouts of immune system genes.
Today I am presenting some interesting findings which have come out recently after several years research.
I will introduce briefly the genetic basis of making a knockout as this is relevant to our findings. Our lab is working on SLE (Systemic Lupus Erythematosus, or Lupus as it is commonly known). This is a multi-system autoimmune disease with a complex genetic basis. Murine models of SLE have proved useful in attempting to identify the genes involved.
Three major mouse models of the disease have been described: NZB/NZW hybrid, MRL lpr and BXSB.
Targeted disruptions of genes of immune system proteins have also been used (gene knockouts) and these seem to have a surprisingly high frequency of autoimmune phenotype associated with them.
Our lab has generated several knockouts of immune system genes.
Today I am presenting some interesting findings which have come out recently after several years research.
I will introduce briefly the genetic basis of making a knockout as this is relevant to our findings.
50. The phenotype of Apcs (SAP) or C1qa deficiency in different mouse strains This is a table showing some of the results from our initial results using two of the gene knockouts generated in our lab: C1q and Apcs. There are distinct strain differences in the amount of autoimmunity developed in these animals after one year. In the Apcs -/- mice the (129 x BL/6) cohort developed high titres of autoantibody against the nucleus (ANA) and considerable autoimmune glomerulonephritis (GN) in the kidney. The C57BL/6 Apcs-/- cohort developed even higher ANA and GN, but the 129Sv cohort did not develop any autoimmunity at all.
In the C1q-/- cohorts, the only strain which developed any autoimmunity was the (129 x BL/6) cohort. Cohorts of C1q -/- in BL/6 and 129Sv were negative.
However the most surprising result for us was the wild type mice in (129 x BL/6), which spontaneously developed autoimmunity!This is a table showing some of the results from our initial results using two of the gene knockouts generated in our lab: C1q and Apcs. There are distinct strain differences in the amount of autoimmunity developed in these animals after one year. In the Apcs -/- mice the (129 x BL/6) cohort developed high titres of autoantibody against the nucleus (ANA) and considerable autoimmune glomerulonephritis (GN) in the kidney. The C57BL/6 Apcs-/- cohort developed even higher ANA and GN, but the 129Sv cohort did not develop any autoimmunity at all.
In the C1q-/- cohorts, the only strain which developed any autoimmunity was the (129 x BL/6) cohort. Cohorts of C1q -/- in BL/6 and 129Sv were negative.
However the most surprising result for us was the wild type mice in (129 x BL/6), which spontaneously developed autoimmunity!
51. Is the hybrid 129 x C57BL/6 genetic background sufficient to induce SLE?
52. Background strain effects on autoimmunity in knockout models of SLE Here I have given a diagrammatic representation of the chromosomes of a wild type (129 x B6) F2 mouse. Due to crossing over the entire genome contains random areas of 129 and B6 DNA. However please note that although the rest of the genome of the Apcs-/- mouse is randomised, chromosome 1 contains an area around the Apcs knocked out gene which is ES cell DNA on both alleles i.e. it is permanently fixed as 129Sv DNA.Here I have given a diagrammatic representation of the chromosomes of a wild type (129 x B6) F2 mouse. Due to crossing over the entire genome contains random areas of 129 and B6 DNA. However please note that although the rest of the genome of the Apcs-/- mouse is randomised, chromosome 1 contains an area around the Apcs knocked out gene which is ES cell DNA on both alleles i.e. it is permanently fixed as 129Sv DNA.
53. Unsorted data for ANA at 12 months of age. A significant statistical difference is found between wild type controls and Apcs-/- cohorts. (Each small dot is a single mouse, the large dot is a group of mice, the number of which is in brackets alongside) Unsorted data for ANA at 12 months of age. A significant statistical difference is found between wild type controls and Apcs-/- cohorts. (Each small dot is a single mouse, the large dot is a group of mice, the number of which is in brackets alongside)
54. Background strain effects on autoimmunity in knockout models of SLE Here I have given a diagrammatic representation of the chromosomes of a wild type (129 x B6) F2 mouse. Due to crossing over the entire genome contains random areas of 129 and B6 DNA. However please note that although the rest of the genome of the Apcs-/- mouse is randomised, chromosome 1 contains an area around the Apcs knocked out gene which is ES cell DNA on both alleles i.e. it is permanently fixed as 129Sv DNA.Here I have given a diagrammatic representation of the chromosomes of a wild type (129 x B6) F2 mouse. Due to crossing over the entire genome contains random areas of 129 and B6 DNA. However please note that although the rest of the genome of the Apcs-/- mouse is randomised, chromosome 1 contains an area around the Apcs knocked out gene which is ES cell DNA on both alleles i.e. it is permanently fixed as 129Sv DNA.
55. ANA data wild type controls sorted for fixed 129 interval. The difference disappears!ANA data wild type controls sorted for fixed 129 interval. The difference disappears!
56. QTL ANALYSIS CHROMOSOME 1
57. Background strain effects on autoimmunity- Congenic strains
58. We aged three cohorts for 12 months. One cohort was a pure BL/6 control, one had a non-functional Apcs gene on an otherwise pure BL/6 background containing the 129Sv KO region on distal Chr1, and the third was a wild type congenic mouse containing a similar (slightly larger) 129Sv segment on Chr 1 with the Apcs gene functional.We aged three cohorts for 12 months. One cohort was a pure BL/6 control, one had a non-functional Apcs gene on an otherwise pure BL/6 background containing the 129Sv KO region on distal Chr1, and the third was a wild type congenic mouse containing a similar (slightly larger) 129Sv segment on Chr 1 with the Apcs gene functional.
59. Phenotypes at 12 months of age were analysed. ANA titre showed that the congenic and the Apcs KO cohort were the same! The control BL/6 mice had a much lower number of positives and were clearly significantly different from either of the other two cohorts.Phenotypes at 12 months of age were analysed. ANA titre showed that the congenic and the Apcs KO cohort were the same! The control BL/6 mice had a much lower number of positives and were clearly significantly different from either of the other two cohorts.
60. CHROMOSOME 1 � SLE candidate genes
61. Gene-targeted mice have contributed to characterise effector mechanisms involved in disease pathogenesis
There are important epistatic interactions between genes from 129 and C57BL/6 mice on the development of SLE
The phenotype of knockout mice can be influenced not only by the disruption of the targeted gene but ALSO by their genetic background
Genes located on telomeric portion of chromosome 1 are involved in SLE pathogenesis
62. Thanks to
63. SLE � pathogenesis Disease pathway Putative disease mechanisms
Dysregulation of the immune system
Abnormal antigen clearance
Uncontrolled inflammation
64. Physiological uptake of apoptotic cells
66. 129 SLE loci on chromosome 1
67. C1q-deficient mice -conclusions C1q inhibits the positive selection of B1 B cells and IgM autoantibodies by an intracellular self antigens
C1q contributes to the clearance of dying cells that may explain the association between complement deficiency and SLE
69. GLOMERULONEPHRITIS
71. SLE - complement, anti-DNA and disease activity
72. Antinuclear antibodies
73. Immune complexes and complement are found in inflamed tissues