Material & Methods
A. SVV261 apoptosis rate 43.4% = 2.2-fold increase. SVV264 apoptosis rate 36.0% = 1.8-fold increase. Systematic in vitro evaluation of survivin-directed antisense oligodeoxynucleotides in bladder cancer cells


Material & Methods
E N D
Presentation Transcript
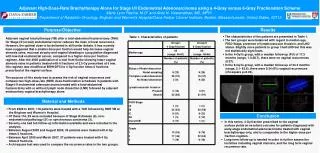
A SVV261 apoptosis rate 43.4% = 2.2-fold increase SVV264 apoptosis rate 36.0% = 1.8-fold increase • Systematic in vitro evaluation of survivin-directed antisense oligodeoxynucleotides • in bladder cancer cells • Susanne Fuessel, Bernd Kueppers, Shuangli Ning, Matthias Kotzsch#, Kai Kraemer, Uta Schmidt, Axel Meye, and Manfred P. Wirth • Department of Urology and Institute of Pathology#, Technical University of Dresden, Germany • Deutsche Krebshilfe e.V. (grant 10-1752) 270 G SVV 261 – 280 SVV 264 – 283 SVV 286 – 305 U 280 290 G C Color code for ss-conservation : ACGU 100% ss ACGU 0% ss ACGU 10 - 90% ss PI-staining SVV286 apoptosis rate 51.3% = 2.6-fold increase NS-K1 apoptosis rate 19.5% Annexin V-staining FIG.3 Effects of six pre-selected AS‑ODN on EJ28 cells. Influence of AS‑ODN on viability and proliferation of transfected EJ28 cells was determined using the WST‑1 and BrdU assays, respectively (in six parallel experiments). For the cell colony formation assay cells were seeded in triplicate (300 cells per 25cm2 flask) and grown for 9 days. The number of colonies was determined using Giemsa staining. Survivin transcript amounts were determined by quantitative real-time PCR in correlation to the reference gene GAPDH (zmol/amol) in duplicate. Survivin protein levels in relation to the total protein content were measured in duplicate by an ELISA. All measurements were normalized to the NS‑K1 control. Values are presented as mean ± SD as for viability and proliferation or as mean ± mean deviation for the other assays. FIG.1 Detail of the predicted secondary structure of the survivin mRNA and aligned AS‑ODN. Displayed are a part of the theoretical secondary structure of the survivin mRNA (Acc.No. NM_001168, energy level (G= -609.8kcal/mol) and three out of 30 tested AS‑ODN hybridizing to their target regions within the mRNA (nt‑positions 261-280, 264-283 and 286-305). Nucleotides are color-coded for ss‑probability within the ten computed energy levels. FIG.5 Effect of AS‑ODN treatment on four different bladder cancer cell lines. Reduction of viability in four BCa cell lines 24 h after the start of transfection with 250 nM of the most effective AS‑ODN tested by WST‑1 assay. Effects on cell growth were normalized to treatment with NS‑K1 (sixfold, mean ± SD). Significance of differences between AS‑ODN and control treatment was calculated bythe unpaired, homoscedastic t‑test (* p<0.05, ** p<0.01, *** p<0.001). Tab.1Sequences of the six most effective anti-survivin AS‑ODN ODN * Sequence # SVV232 § cccagccttccagctccttg SVV261 atgttcctctatggggtcgt SVV264 tttatgttcctctatggggt Tab.2 Cell cycle alterations in EJ28 Cells 24 h after AS‑ODN treatment SVV286 aagcgcaaccggacgaatgc FIG.2 Survivin expression in different BCa cell lines. Survivin transcript levels [zmol] in different BCa cell lines and in human foreskin fibroblasts (FB) were determined by quantitative real-time PCR and normalized to the mRNA expression of the reference gene GAPDH [amol]. Survivin protein expression was measured in the same cell lines using an ELISA and normalized to the total protein content. SVV532 gctggtggcaccagggaata ODN G0 / G1 (%) S (%) G2 / M (%) SVV743 acctggtaagcccgggaatc FIG.4 Induction of apoptosis in EJ28 cells 24 h after AS‑ODN treatment. Apoptosis rates in EJ28 cells were measured by FACS analysis after Annexin V-FITC staining. PI counterstaining allowed the differentiation between viable cells (lower left quadrant), early apoptotic cells (lower right quadrant), late apoptotic cells (upper right quadrant) and necrotic cells (upper left quadrant). The percentages of early and late apoptosis representing the total amount of apoptotic cells were summed up. The increase of apoptosis was calculated in relation to the NS‑K1 control. NS-K1 $ taagctgttctatgtgtt SVV261 53.2 31.7 15.1 * Designation of AS‑ODN derived from starting position within the survivin mRNA sequence (Acc.No. NM_001168). Less effective AS‑ODN were targeted at the following starting nt‑positions within the survivin mRNA: 33, 41, 49, 278, 282, 501, 504, 516, 519, 526, 716, 719, 724, 740, 1099,1101, 1103, 1104, 1126, 1128, 1302, 1304, 1317, 1325 and 1327. # All AS‑ODN were 20 nt in length (except SVV1099 and NS‑K1) and protected by two PTO, each at the 3’-end and at the 5’-end. § deduced from construct AS4003 described by Olie et al. $ deduced from construct MS‑ODN described by Chen et al. SVV264 48.6 36.4 15.0 SVV286 53.3 37.4 9.3 NS‑K1 44.5 43.8 11.7 Objectives The rather poor responses to conventional treatment of bladder cancer (BCa) require novel, specific therapy approaches. The down-regulation of BCa-associated genes may represent a new option to specifically inhibit BCa cell growth and to induce cell death. Survivin, an inhibitor of apoptosis upregulated in the vast majority of malignancies including BCa provides an attractive target for molecular therapies such as treatment with specific antisense oligodeoxynucleotides (AS‑ODN). Results Survivin mRNA and protein expression was analyzed in eight BCa cell lines by quantitative RT‑PCR and ELISA. All investigated BCa cell lines showed moderate to strong survivin expression (Fig.2). The cell line EJ28 displaying the highest survivin levels (Fig.2) was selected for primary screening. In a second step, the cell lines 5637, J82 and HT1197 were also investigated to test the efficacy of the initially selected AS‑ODN. Initially, the inhibitory efficacy of all 30 designed AS‑ODN was analyzed by measurements of the viability of EJ28 cells. Six AS‑ODN (SVV232, SVV261, SVV264, SVV286, SVV532 and SVV743) with the highest inhibitory effects were selected for more detailed studies (Fig.3). The viability (WST‑1 assay) and the proliferation (BrdU assay) of EJ28 cells were reduced to 40-60% by the selected constructs as compared to the control NS‑K1 (Fig.3). Additionally, a considerable effect with inhibition rates between 48% (SVV232) and 86% (SVV261) on clonogenic survival of EJ28 cells after AS-ODN transfection was observed in the cell colony formation assay (Fig.3). Simultaneously, these treated EJ28 cells showed a clear reduction of survivin transcript and protein levels of up to 60-80% in relation to NS-K1 (Fig.3). In cell cycle analyses, a reduction in the percentage of cells in S‑phase in favor of cells arrested in G0/G1 and/or G2/M was observed for the AS‑ODN SVV261, SVV264 and SVV286 (Tab.2). Measurements of apoptosis (Fig.4) showed that AS‑ODN SVV286 caused a 2.6-fold increase of apoptosis in relation to NS‑K1 followed by SVV261 (2.2-fold) and SVV264 (1.8-fold). To validate the effects as assessed for EJ28 cells, we investigated the influence of the AS‑ODN SVV261, SVV264 and SVV286 on the viability of further BCa cell lines (5637, HT1197 and J82). The reduction of cell growth by these constructs could be confirmed, although the degree of inhibition varied between the different BCa lines (Fig.5). In 5637 cells expressing survivin at a similar level as EJ28 cells (Fig.2), a comparable inhibition of viability was observed. A lower but obvious inhibition of viability was shown for the lines HT1197 and J82 possessing lower amounts of survivin transcripts and protein (Fig.2). Material & Methods A total of 30 AS-ODN were designed against 5 single stranded (ss) motifs of the survivin mRNA, predicted by secondary structure calculation (mfold software 3.1; Fig.1). A nonsense ODN (NS-K1) served as control in all experiments (normalization of the effects with NS-K1= 100%). The BCa cell lines EJ28, 5637, J82 and HT1197 were transfected for 4h with 250nM ODN and Lipofectin (ratio 1:3; Invitrogen). To screen for viable cells we employed the WST-1 reagent (Roche). Incorporation of 5‑Bromo‑2’‑deoxy-uridine (BrdU) was assessed to measure the cellular proliferation (colorimetric BrdU Cell Proliferation ELISA, Roche). In order to test the colony formation, 300 cells were plated in 25cm2-flasks and incubated for 7 days. After Giemsa staining, the clonogenic survival was determined by counting all macroscopically visible colonies. Cell cycle analysis was performed with the CycleTest Plus DNA Reagent Kit (BD Biosciences) using flow cytometry. Rates of apoptosis were assessed by flow cytometry after double-staining cells with FITC-labeled AnnexinV (AnnexinV-FITC Apoptosis Detection Kit, BD) and propidium iodide (PI). Total cellular RNA was isolated by a standard protocol (Invisorb Spin Cell RNA Mini Kit, Invitek, Berlin, Germany) 24 h after the start of transfection. The survivin mRNA was quantified by real time RT-PCR. Survivin transcript amounts were normalized to mRNA levels of GAPDH measured within the same sample. These relative mRNA expression levels (zmol survivin/ amol GAPDH) were compared to those in cells treated with NS-K1. Survivin protein was determined quantitatively in lysates of cells harvested 24 h after the start of transfection using the survivin DuoSet IC-ELISA (R&D Systems) with some modifications. Survivin protein levels were normalized to total protein content and expressed as ng per mg protein. Conclusions Three of 30 tested AS-ODN reduced the growth of BCa cells specifically. These efficient AS-ODN caused a noticeable reduction in survivin expression. Interestingly, all three efficient constructs were directed towards nt‑position 274-285. This putative ss‑region appears to be qualified as mRNA target motif since other nucleic acid based inhibitors like ribozymes [Choi et al.] or siRNA constructs [Williams et al.] targeted at the same motif were reported to be highly active. The survivin-directed AS‑ODN shown to inhibit the expression of this inhibitor of apoptosis and to impair subsequently the proliferation of BCa cells in the present study, may provide suitable adjuvant therapeutic agents for the specific local treatment of BCa. References Chen, J., Wu, W., Tahir, S.K., Kroeger, P.E., Rosenberg, S.H., Cowsert, L.M. et al. (2000) Neoplasia, 2: 235-41. Olie, R.A., Simoes-Wust, A.P., Baumann, B., Leech, S.H., Fabbro, D., Stahel, R.A., and Zangemeister-Wittke, U. (2000) Cancer Res, 60: 2805-9. Choi, K.S., Lee, T.H., and Jung, M.H. (2003) Cancer Gene Ther, 10: 87-95. Williams, N.S., Gaynor, R.B., Scoggin, S., Verma, U., Gokaslan, T. et al. (2003) Clin Cancer Res, 9: 931-46. www.tu-dresden.de/meduro susanne.fuessel@mailbox.tu-dresden.de