Download

1 / 31
310 likes | 620 Vues
Mekanisme Molekuler Induksi Tumor Crown Gall oleh Agrobacterium tumefaciens. III. DISTRIBUSI SEBUAH GEN VIRULEN KROMOSOM, acvB , DARI AGROBACTERIUM TUMEFACIENS PADA BERBAGAI BAKTERI. TIK : Mahasiswa mengetahui teknik distribusi gen virulen kromosom pada bakteri.

E N D
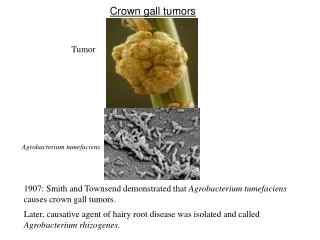
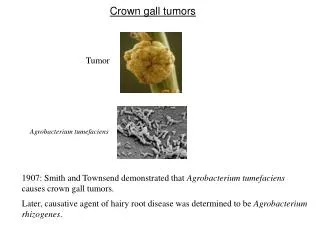
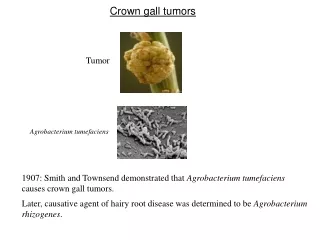
Mekanisme Molekuler Induksi Tumor Crown Gall oleh Agrobacterium tumefaciens
III. DISTRIBUSI SEBUAH GEN VIRULEN KROMOSOM, acvB, DARI AGROBACTERIUMTUMEFACIENS PADA BERBAGAI BAKTERI TIK : • Mahasiswa mengetahui teknik distribusi gen virulen kromosom pada bakteri. • Mahasiswa mampu menganalisa hasil distribusinya gen virulen kromosom pada bakteri. Waktu: 2x(2x50 menit)
Sub Pokok Bahasan: 3.1 Pendahuluan 3.2 Bahan dan Teknik Distribusi Gen Virulen KromosomPada Bakteri 3.2.1 Strain Bakteri, Plasmid, dan Kondisi Kultur 3.2.2 Pembuatan Probe DNA [32P] 3.2.2 Hibridisasi Southern 3.2.3 Pembuatan Rekombinan Pertukaran Marker 3.2.4 Pengujian Virulensi 3.2.5 Fraksinasi Sel Agrobacterium 3.2.6 Pengekspresian gen acvB dan lokalisasi selular dari produknya dalam berbagaibakteri
3.3 Analisa Hasil Distribusi Gen Virulen Kromosom padaBakteri 3.3.1 Pengujian Keberadaan gen acvB dalam berbagai bakteri 3.3.2 Pengaruh perusakan gen acvB kromosomal pada virulensi A.tumefaciens yang memiliki Ti plasmid tipe oktopin dan nopalin. 3.3.3 Pengujian Open Reading Frame dari Gen acvB oleh program Analisis "Frame". 3.3.4 Analisi Komplementari. 3.4 Diskusi dan Kesimpulan
3.1 PENDAHULUAN • Pada bab sebelumnya, telah digambarkan isolasi dua mutan avirulen (B90 dan B119) dan dua gen virulen kromosom (acvI dan acvB) dari mutan-mutan tersebut. • Kedua mutan dan kedua gen tersebut berbeda dari mutan-mutan dan gen-gen kromosom yang dilaporkan sebelumnya. • Sebagai tahap penelitian berikutnya, perlu diuraikan pada proses manakah dan bagaimanakah dalam induksi Crown gall gen-gen virulen ini berfungsi. Pada pokok bahasan ini akan difokuskan pada satu dari dua gen di atas, yaitu acvB.
Gen acvB adalah gen virulen kromosom baru. Penelitian homologinya mengungkapkan bahwa daerah C-terminal dari AcvB menunjukkan sebuah kesamaan yang signifikan dengan N-terminal pada bagian hilir (downstream) ORF dari gen virA pada Ti-plasmid tipe-oktopin (45, 88). Sayangnya, fungsi dari ORF masih belum diketahui. • Dalam bab ini, dibahas tentang distribusi gen acvB di antara berbagai macam bakteri sebagai langkah awal penelitian tentang fungsi dari gen acvB.
3.2 BAHAN DAN TEKNIK DISTRIBUSI GEN VIRULEN KROMOSOM PADA BAKTERI 3.2.1 Strain Bakteri, Plasmid, dan Kondisi Kultur • Bakteri dan plasmid yang digunakan pada bab ini disusun dalam Tabel 2. Agrobacterium ditumbuhkan pada medium minimal AB (13) pada suhu 28°C. • Escherichia coli, pseudomonas fluorescens dan Bacillus subtilis ditumbuhkan pada cawan agar L dan di dalam L broth pada suhu 37°C dan Rhizobia pada suhu 28°C.
Konsentrasi (µg/ml) antibiotik yang digunakan di dalam media adalah sebagai berikut: untuk Agrobacterium tumefaciens, kanamisin, 100; ampicilin, 50; gentamisin, 100; tetracylin, 10. Untuk E. Coli, kanamisin, 50; ampisilin, 100; tetrasiklin, 20; gentamisin 100. • Masuknya plasmid ke dalam Agrobacterium dilakukan dengan perkawinan tiga induk (triparental mating) menggunakan plasmid pembantu (helper plasmid), pRK2013, seperti yang disebutkan sebelumnya (20).
3.2.2 Pembuatan Probe DNA [32P] • Probe 1 disiapkan dengan cara pemberian label fragmen Ava I-EcoR I (500 bp) pada ujung 3’ dari gen acvB dengan menggunakan [α-32P]-dCTP dan sebuah sistem pelabelan primer acak (Takara). • Probe 2 diberi label 32P pada fragmen IS50 dari Tn5 yang dibuat menggunakan [α-32P]dCTP dan sistem pelabelan primer acak.
3.2.3 Hibridisasi Southern • DNA total diekstrak dengan metode Kado dan Liu (41) dari berbagai bakteri termasuk Agrobacterium, Rhizobia, Psedomonas, dan Bacillus. • Jumlah yang sama (1µg) dari DNA total digunakan pada elektroporesis gel agarose 0.8% untuk memisahkan plasmid dan DNA kromosom. • Gel dikeringkan di atas membran nilon Hybond N+ (Amersham) dan dihibridisasi dengan probe 1. • DNA Kromosom (1µg) dari strain-strain B119, A208ME dan A348ME dicerna dengan Pst I, BamH I, atau EcoR I, dielektroporesis dengan gel agarose (1%), dikeringkan di atas sebuah membran nilon dan dihibridisasi dengan probe 2. • Pita (band) yang telah dihibridisasi diamati dengan penganalisa gambar (Bas 2000, Fujix).
3.2.4 Pembuatan Rekombinan Pertukaran Marker • Sebuah plasmid pNU119 yang mengandung gen acvB dengan sisipan Tn5 pada bagian tengahnya, dipindahkan ke dalam A. tumefaciens (strain A208) yang mengandung Ti plasmid tipe napolin, dan A. tumefaciens (strain A348) yang mengandung Ti plasmid tipe oktopin, dengan konjugasi perkawinan tiga induk (triparental mating conjugation) (83) menggunakan E. Coli yang membawa plasmid pembantu (helper) pRK2013 (20). • Kemudian dipilih transkonjugan yang resisten terhadap tetrasilin dan kanamisin yang tumbuh pada medium minimal AB. Plasmid yang dikeluarkan, pPh1J1 (32), dipindahkan ke dalam strain yang dihasilkan, dan koloni yang resisten terhadap kanamisin dan gentamisin dipilih pada medium AB.
Marker exchange recombinants dari kedua strain Agrobacterium diisolasi dengan melakukan screening koloni yang resisten terhadap kanamisin dan gentamisin untuk sensitivitas terhadap tetrasiklin. • Percobaan yang sama juga dilakukan pada A.tumefaciens (strain ACTT23308) dan A. rhizogeneses (Strain IFO 13259).
3.2.5 Uji Tingkat Virulensi • Rekombinan Pertukaran Marker, yaitu strain A208ME dan A348ME serta strain-strain induk, yaitu strain A208 dan A348, diinokulasi pada Kalanchoadiagremontiana untuk diuji tingkat virulensinya. • Foto diambil dua bulan setelah inokulasi dilakukan.
3.2.6 Fraksinasi Sel pada Agrobacterium • Fraksinasi pada sel Agrobacterium dilakukan sesuai dengan metode yang telah dilaporkan sebelumnya dengan sedikit modifikasi (19). • Sel tersebut ditumbuhkan selama semalam dalam medium cair YEB (40ml) pada suhu 28 °C dan dikumpulkan dengan sentrifugasi (10 k x g selama 5 menit). • Peletnya disuspensi dalam 5 ml Tris-HCL 50mM, pH 8,0, yang mengandung sukrosa (20%), EDTA (2mM), serta lisozim (0.2mg/ml) dan diinkubasi pada suhu 28°C selama 30 menit dengan pengocokan perlahan. • Kemudian suspensi sel disentrifugasi pada 10 x g selama 5 menit pada suhu 4°C untuk memisahkan supernatan dan peletnya (10 k x g - pelet).
Protein periplasma dipisahkan dari supernatan, kemudian protein-protein membran dan sitoplasma disiapkan dari pelet. • Asam trikloroasetat (TCA) ditambahkan pada supernatan untuk mendapatkan konsentrasi akhir 5%(w/v) dan disimpan dalam es/pendinginan selama 30 menit. • Endapan yang terbentuk dipisahkan dengan sentrifugasi, dicuci dua kali dengan dietil eter dan dilarutkan dalam Tris-HCl 50 mM, pH 8.0 (protein periplasma). • Pellet 10 k x g disuspensi kembali dalam 5 ml Tris-HCl 50 mM, pH 8.0, dan disonikasi dengan singkat untuk membelah selnya.
Kemudian, suspensi tersebut disentrifugasi pada 12 k x g selama 5 menit untuk memisahkan sel yang tidak terbelah. • Supernatan yang dihasilkan disentrifugasi pada 213 k x g selama satu jam pada suhu 4°C untuk memisahkan supernatan dari endapannya (protein membran). • TCA (5%, w/v) ditambahkan ke dalam supernatan dan disimpan dalam es/pendinginan selama 30 menit. • Endapan yang terbentuk dipisahkan dengan sentrifugasi (12 k x g selama 15 menit), dicuci dengan dietil eter, dan dilarutkan dalam Tris-HCl 50 mM, pH 8.0 (protein sitoplasma). • Fraksinasi sel bakteri lainnya dilakukan dengan menggunakan prosedur yang sama seperti prosedur yang dipakai pada Agrobacterium.
Untuk menguji kemurnian fraksi yang telah dipisahkan, maka alkalin, pospatase, NADH oksidase, dan malate dehidrogenase dari fraksi tersebut diuji kadarnya sebagai sebuah enzim penanda fraksi periplasma, membran, dan sitoplasma, secara berturut-turut, dengan metode yang telah dilaporkan (19). • Satu unit aktivitas enzim pada masing-masing enzim didefinisikan sebagai aktivitas yang memproduksi perubahan sebesar 0.001 kerapatan optik pada panjang gelombang yang diukur per menit di bawah kondisi pengujian kadar.
3.2.7 Ekspresi gen acvB dan Lokalisasi Seluler Produknya pada Beberapa Bakteri • Sel-sel dari bakteri gram negatif termasuk A.tumefaciens (strain A208 dan A348), A.rhizogenes, A.radiobacter, Rhizobium meliloti, R.Phaseoli, dan Pseudomonas fluorescens difraksinasi menjadi fraksi periplasma, sitoplasma, dan membran sesuai dengan prosedur yang digambarkan sebelumnya. • Bacilluss subtilis (gram positif) juga difraksinasi dengan metode yang sama. • Potein (12 µg) dari fraksi-fraksi tersebut dipisahkan dengan SDS-PAGE, dikeringkan secara elektronik (electro-blotted) diatas sebuah membran ImobilonTMPVDF (Millipore) pada 40 mA selama satu jam dan diwarnai secara imunologi (immuno-stained) menggunakan serum anti-AcvB seperti yang dilaporkan (69).
3.3. ANALISA HASIL DISTRIBUSI GEN VIRULEN KROMOSOM PADA BAKTERI 3.3.1 Pengujian Keberadaan Gen acvB pada Berbagai Bakteri • DNA total diekstrak dengan metode Kado and Liu (41) dari beberapa bakteri termasuk strain Agrobacterium, Rhizobium, Pseudomona dan Bacillus. Jumlah yang sama (1µg) dari DNA total digunakan untuk elektroporesis gel agarose 0.8% untuk memisahkan plasmid dan DNA kromosom. • Gel tersebut dikeringkan di atas membran nilon dan dihibridisasi dengan fragmen DNA acvB berlabel 32P (probe 1) (Gb. 16).
Gen acvB dideteksi pada semua strain Agrobacterium yang diuji; A.tumefaciens (strain A208) yang memiliki sebuah Ti plasmid tipe nopalin pTiT37 (lane 1), A.rhizogenes (strain IFO13259) (lane 2), A.radiobacter (strain IFO 12665) (lane 3), A.tumefaciens (strain ACTT23308) yang memiliki Ti Plasmid tipe oktopin (lane 4), dan A.tumefaciens (strain A348) yang memiliki Ti plasmid tipe oktopin pTiA6 (lane 5). • Pada strain A.tumefaciens yang memiliki Ti plasmid tipe oktopin, gen acvB dideteksi baik pada Ti plasmid maupun kromosomnya (jalur 4 dan 5), sementara gen acvB hanya dideteksi pada DNA kromosom dalam A.tumefaciens yang memiliki Ti plasmid tipe napolin (lane 1). • Bakteri lain selain Agrobacterium tidak mengandung gen acvB.
3.3.2 Efek Gangguan Gen kromosom acvB pada virulensi A.tumefaciens yang memiliki Ti plasmid tipe oktopin atau nopalin • Untuk menguji apakah gen homolog acvB pada Ti plasmid tipe oktopin berfungsi atau tidak, maka dilakukan percobaan gangguan gen. • Sebuah plasmid pNU119 yang mengandung gen acvB dengan sisipan Tn5 pada bagian tengahnya, dipindahkan ke dalam A.tumefaciens (strain A208) yang memiliki Ti plasmid tipe nopalin dan A.tumefaciens (strain A348) yang memiliki Ti Plasmid tipe oktopin untuk mengisolasi rekombinan pertukaran marker pada kedua strain. • Tn5 diletakkan hanya pada DNA kromosom namun tidak pada Ti plasmid dalam kedua rekombinan (data tidak ditunjukan). • Lebih lanjut, DNA-DNA kromosom dari kedua rekombinan dan strain B119 menunjukkan profil fragmen DNA yang tepat sama ketika DNA-DNA tersebut direaksikan dengan tiga jenis enzim restriksi, PstI, BamHI, dan EcoRI, dan dianalisis dengan hibridisasi Southern menggunakan IS50 berlabel-[32P] dari Tn5 (probe 2) (Gb. 17).
Hasilnya menunjukkan bahwa rekombinasi yang sama terjadi pada tempat yang sama dari DNA kromosom pada kedua rekombinan dengan pada penyisipan Tn5 dalam strain B119. • Rekombinan-rekombinan tersebut diinokulasi pada Kalanchoe diagremontiana untuk menguji virulensi (sifat racunnya) (Gb. 18A). • Rekombinan pertukaran marker (A208ME) bersifat avirulen. Sebaliknya, rekombinan pertukaran marker (A348ME) bersifat virulen meskipun gen acvB pada kromosomnya diganti dengan gen acvB yang termutasi.
Percobaan gangguan gen juga dilakukan pada A.tumefaciens (strain ACTT23308) yang memiliki Ti plasmid tipe oktopin dan A.rhizogeneses (strain IFO13259). • Rekombinan A.tumefaciens (strain ACTT23308) yang memiliki gen acvB terganggu pada kromosomnya bersifat virulen, sementara rekombinan dari A. Rhizigenes (strain IFO13259) yang memiliki gen acvB terganggu pada kromosomnya bersifat avirulen (Gb. 18B dan Gb. 18C). • Dengan demikian, hal ini menunjukkan bahwa kesamaan acvB (acvBhomolog) pada Ti plasmid tipe oktopin berfungsi dalam A.tumifaciens.
3.3.3 Ekspresi Gen acvB dan Lokalisasi Seluler dari Produk Gen-nya pada Berbagai Bakteri. • Analisis Western Blot dilakukan untuk menguji ekspresi gen acvB dan lokalisasi dari produk gen-nya pada berbagai sel bakteri. • Sel bakteri difraksinasi menjadi fraksi periplasmik, sitoplasmik, dan membran sesuai dengan prosedur yang digambarkan sebelumnya. • Kemurnian fraksi-fraksi tersebut diuji dengan mengukur tingkat aktivitas enzim penanda (marker enzymes) dari masing-masing fraksi; alkalin pospatase (periplasma), NADH oksidase (membran), dan malat dehidrogenase (sitoplasma) (Tabel 3). • Sembilan puluh tiga % dari total alkalin pospatase terletak pada fraksi periplasmik. Malat dehidrogenese sebagian besar ada pada fraksi sitoplasmik (64%), sementara sebagian besar NADH oksidase berada pada fraksi membran (82%).
Distribusi enzim penanda yang sedemikian rupa diantara fraksi-fraksi menunjukkan bahwa setiap fraksi yang terpisah tidak terkontaminasi-silang dengan parah. • Protein dari fraksi-fraksi tersebut dipisahkan dengan SDS-PAGE, ditempelkan secara elektronik (elektro-blotted) pada membran PVDF dan diwarnai secara imunologi (immuno-stain)menggunakan serum anti-AcvB. • Seperti yang ditunjukan Gb. 19, protein AcvB dideteksi di dalam fraksi periplasma dari A.tumefaciens (strainA208) (lane 2), A.tumefaciens (strain A348) (lane 3), A.tumefaciens (strain ACTT23308) (lane 4), A.rhizogenes (strain IFO13256) (lane5) dan A.radiobacter (strain IFO12665) (lane 6). • Namun, fraksi-fraksi sitoplasma dan membran dari bakteri-bakteri ini tidak mengandung protein AcvB (data tidak ditunjukkan). Selain itu, protein AcvB tidak terdeteksi pada belahan R.meliloti (lane 7), R.phaseoli (lane 8), P.fluorescens (lane 9) atau B.subtilis (lane 10). Dengan demikian, semua bakteri yang mengandung gen acvB mengekspresi protein AcvB yang terletak dalam periplasmanya.
3.4 DISKUSI DAN KESIMPULAN • Dalam strain A.tumefacien yang memiliki Ti plasmid tipe oktopin, gen acvB dideteksi pada Ti plasmid dan kromosomnya, sementara gen acvB hanya dideteksi pada DNA kromosom dalam A.tumefaciens yang memiliki Ti plasmid tipe napolin (Gb. 16). • Hasil ini sesuai dengan fakta dimana pencarian homologi menggunakan E-mail server NCBI Gen Bank mengungkapkan bahwa homologi gen acvB hanya ada dengan susunan di antara gen virA dan virB pada Ti Plasmid tipe oktopin tetapi tidak terdapat dalam Ti plasmid tipe napolin (45, 88).
Di samping itu, hasil ini sesuai dengan paper terbaru yang menjelaskan tentang terbentuknya sebuah gen (virJ) pada Ti plasmid tipe oktopin (43) yang homolog dengan C-terminal setengah dari gen acvB. • Hasil dari percobaan gangguan gen pada Gb. 18 menunjukkan bahwa homolog gen acvB (virJ) berfungsi dalam A.tumefaciens. • Hasil ini sesuai dengan hasil yang diperoleh oleh V.S. Kalogeraki dan S.C.Winans (43) bahwa virJ dapat melengkapi penotip avirulen dari strain mutan B119 (acvB-).
Dari semua strain Agrobacterium yang diuji dalam studi ini, terdapat gen acvB. Sebaliknya, gen acvB tidak ditemukan dalam bakteri lainnya. • Agrobacterium mampu mentransfer DNA ke dalam sel tanaman, sementara bakteri lain yang diuji dalam studi ini tidak mampu mentransfer DNA ke dalam sel tanaman. • Akibatnya, distribusi gen acvB yang seperti ini menunjukkan bahwa gen acvB terlibat dalam proses transfer DNA pada sel tanaman.
Agrobacteria dan Rhizobia memiliki kesamaan dalam banyak aspek; keduanya merupakan bakteri gram negatif dari tanah (soil born) yang mengandung Ti atau Sym plasmid, secara berurutan. • Strain Agrobacterium menginduksi Crown gall dan strain Rhizobium menginduksi bintil-bintil akar (nodules) pada tanaman inang. Kedua bakteri ini sama-sama mengandung banyak kesamaan gen. Contohnya, gen chvA dan chvB yang sama ditemukan pada kedua bakteri ini (8,23), dan kedua gen ini terlibat dalam sintesis dan transpor β-1,2-glucan pada kedua bakteri tersebut. • Walaupun demikian, gen acvB hanya terdapat dalam strain Agrobacterium namun tidak pada strain Rhizobium
Meskipun Agrobacteria dan Rhizobia memiliki kesamaan yang ekstensif, terdapat perbedaan besar dalam mekanisme yang besar dalam proses induksi crown gall oleh Agrobacterium dan induksi bintil-bintil akar (nodule) oleh Rhizobia. • Selama induksi Crown gall oleh Agrobacterium, T-DNA ditransfer di dalam (inside) sel tanaman, sementara DNA tidak ditransfer di dalam sel tanaman selama induksi bintil-bintil akar oleh Rhizobium.
Dengan demikian, proses transfer DNA ke dalam sel tanaman merupakan proses yang hanya berlangsung pada Agrobacterium namun tidak pada Rhizobium. Sebagai hasilnya, acvB kemungkinan terlibat dalam proses ini. • Kesimpulannya, hasil yang diperoleh dari studi ini menunjukkan bahwa gen acvB terlibat dalam proses unik yang hanya terjadi pada Agrobacterium, yakni proses transfer T-DNA dari Agrobacterium ke sel tanaman inang.