Download

1 / 76
760 likes | 801 Vues
Dive into RNA processing, DNA synthesis, and chromatin control in this comprehensive exploration of molecular biology methods and techniques. Discover the intricacies of transcription regulation and post-transcriptional modifications.

E N D
Molecular Biology:Translation and Methods Andy HowardBiochemistry Lectures, Spring 2019Tuesday 12 March 2019
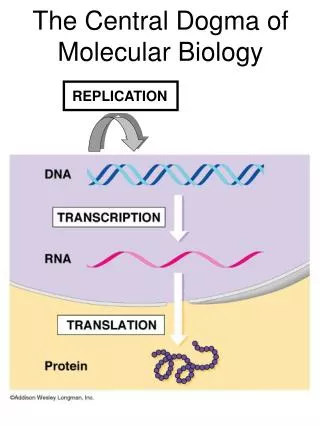
DNA can be manipulated • We’ll show how DNA is replicated in the laboratory and we’ll explore some specific aspects of recombinant DNA technology and its applications • But first we’ll finish talking about protein synthesis and DNA reactivity Translation; Mol Bio Methods
What we’ll discuss • RNA processing • Translation • Tertiary Structure • DNA winding and gyrases • DNA synthesis • Cloning • Probes • Hybridization, blots • High-throughput:Microarrays • Prokaryotic Expression • CRISPR / Cas9 Translation; Mol Bio Methods
Chromatin’s role • A huge percentage of genomic DNA remains untranscribed within any given cell • Most of the decision-making occurs at the level of the chromatin complexes in the nucleosome • Histones: acetylations, deacetylations, methylations of K; methylations of R; phosphorylations of S & T are signals • Nucleosome positioning & remodeling Translation; Mol Bio Methods
How is chromatin controlled? J.Lindsay et al. (2005) Neurosurg. Focus19:5 • Protein complexes use ATP hydrolysis to remodel the nucleosomes and provide access of proteins to the DNA • Some of the complexes contain HATs or HDACs Translation; Mol Bio Methods
Regulation of transcription • A few genes are transcribed richly in every cell: they have strong promoters • Many mechanisms for regulating the rest • Both repressors and activators exist • Typically allosteric proteins whose functions are influenced by ligand binding • For genes that code for metabolic enzymes, the cell can control transcription to make available the appropriate proteins only when it’s useful Translation; Mol Bio Methods
Repression examples • Example 1: repressor for metabolic genes prevents transcription in absence of substrate; when substrate (“inducer”) binds to repressor, it stops repression and allows transcription; this means no needless transcription • Example 2: repressor operates only when downstream metabolite (“corepressor”) binds to repressor; this prevents overproduction on pathway Translation; Mol Bio Methods
β-galactosidase processing E.coli doesn’t ordinarily transcribe the genes required for lactose processing in high quantities If it needs to rely on β-galactosides, it will Three proteins required: Translation; Mol Bio Methods
The lac operon E.colilac repressor 155 kDa tetramer PDB 3EDC, 2.1Å These 3 genes: transcribed from singlepromoter to produce 1 big mRNA Homotetrameric regulatory protein called lac repressor coded for by lacI Repressor binds to two “operator” sites near promoter: O1 next to promoter, O2 within coding region of lacZ Translation; Mol Bio Methods
Ka(O1) = 1013M-1 Kinetics of repression • When repressor binds to both operators, it pushes the DNA into a stable loop • Either prevents binding of polymerase or prevents conversion of closed complex to open complex • Repressor binds nonspecifically; slides until it finds the operators Translation; Mol Bio Methods
lactose Inducers β-galacto-sidase allolactose Various β-galactosides can bind repressor and reduce its affinity for the operators 104-fold Allolactose (produced by action of β-galactosidase on lactose) is a potent inducer Translation; Mol Bio Methods
Regulation by cAMP E.coli CRP mutant48 kDa homodimerPDB 1HW5, 1.8Å • Cyclic AMP regulatory protein (CRP) has low affinity for DNA in absence of cAMP • With cAMP it becomes an activator that binds just upstream of -35 region and increases transcription • [cAMP] controlled by reductions in extracellular [glucose] via phosphate activation of adenylate cyclase Translation; Mol Bio Methods
Post-transcriptional modifications of RNA ThermatogaRNase P RNA & protein + tRNA157kDa total MWPDB 3Q1R, 4.21Å • Every form of RNA can be modified after transcription • tRNA: initial transcript contains several tRNA precursors • RNAse P breaks that into pieces • RNase D trims 3’ end • tRNAnucleotidyl transferase adds CCA to 3’ end • Covalent modifications of bases Translation; Mol Bio Methods
Modifications of rRNA AquifexRNase III+dsRNA 66.8 kDa total PDB 2NUG, 1.7Å • Primary transcript in prokaryotes=30S: one copy each of 16S, 23S, 5S rRNAs, plus interspersed tRNAs • 5’, 3’ ends of each mature rRNA contain base-paired regions in primary transcript • RNase III binds & cleaves; then further trimming • Eukaryotic processing similar; processed in nucleolus Translation; Mol Bio Methods
Eukaryotic mRNA processing • Modifications increase stability to exonucleases • 5’ end modified during synthesis in nucleus • triphosphate loses one phosphate; • XDP reacts with GTP: forms triphosphate + PPi: the “cap” • then the capping guanine gets methylated • CPSF added, polyA polymerase binds, endonuclease acts, polyA tail added Translation; Mol Bio Methods
Splicing • Internal sequences (introns) in messageremoved; remaining pieces are exons • Junctions between those are splice sites • 5’ splice site: GURAGU • 3’ splice site (branch site): YNYURAY • Probably introns were inserted into genes relatively late in evolution; therefore: little evidence of correlations between structural domains and exons Translation; Mol Bio Methods
In the spliceosome Human Dim2 34kDa dimerPDB 3GIX, 1.33Å • Transesterification: 5’splice site to branch-site adenylate:makes lariat-shaped intron • Transesterification between 5’ exonand 3’ splice site (becomes joined exon) • Spliceosome keeps products positioned: • > 100 proteins • 5 snRNA molecules (~5000 nucleotides) • Each snRNA has proteins associated with it: snRNPs; these are used for other things too • 5 kinds of snRNAs: U1, U2, U4-6. Translation; Mol Bio Methods
Probable sequence of events • Rate-limiting step is intron removal Translation; Mol Bio Methods
iClicker question #1 1. Why is repair more important in replication than in transcription? • (a) RNA is less prone to mispairing than DNA • (b) transcription is slow, so it’s less likely to incorporate errors • (c) the errors don’t matter when they happen in the introns • (d) Errors in DNA are persistent; errors in RNA are limited to the immediate transcript. Translation; Mol Bio Methods
Protein synthesis: overview • Proteins are synthesized at the ribosome, a molecular machine found in cytosol and in some organelles • Each amino acid is added to a growing protein chain from the N-terminal end toward the C-terminal end • Each activated amino acid is delivered by a tRNA molecule to the ribosomal machinery • Actual synthesis involves initiation, elongation, termination Translation; Mol Bio Methods
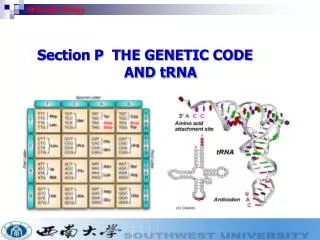
The genetic code Marshall Nirenberg George Gamow • Defined as the relationshipbetween genetic information and the amino acid sequences produced by it • Gamow suggested that three-base codes were the most likely (4 bases, 20 aa’s): • Two insufficient (42 = 16 < 20) • Four excessive (44 = 256 >> 20) • Overlapping code is possible, but it turns out that the real one is non-overlapping Translation; Mol Bio Methods
Features of the code H. Gobind Khorana • Each codon codes for exactly one aa • Code is degenerate, i.e. multiple 3-base codons can code for the same amino acid • First 2 bases of the codon often enough • Similar sequences=>similar aa’s, especially at the middle base • 3 of 64 codons specify stop codons • AUG (met) also specifies initiation site Translation; Mol Bio Methods
The code Translation; Mol Bio Methods
Transfer RNA Sketch courtesy UIC Characteristic cloverleaf structurecontaining Watson-Crick base-pairs 5’ and 3’ ends base-paired to each other Amino acid attached to A at acceptor stem (3’ end) Single-stranded region opposite acceptor stem is anticodon, i.e. the three-base sequence complementary to mRNA codon; it binds that mRNA codon Translation; Mol Bio Methods
tRNA-mRNA base-pairs Normally all 3 bases are base-paired But 3’ and middle bases are the only required ones; 5’ end of anticodon (“wobble base”) can violate strict rules Translation; Mol Bio Methods
Variations on themes Some tRNAs can recognize more than one codon Sometimes we need multiple tRNAs to recognize all synonymous codons Those are called isoacceptor tRNAs, which also applies to tRNAs with same anticodon, different primary sequences Bacterial genomes: 30-60 tRNAs;eukaryotes ~ 80 tRNAs, multiple copies Translation; Mol Bio Methods
Aminoacyl-tRNA synthetases E.coli met-tRNAsynthetaseEC 6.1.1.1063 kDa monomerPDB 3H9C, 1.4Å • Enzymes:catalyze covalent attachmentof an amino acid to 3’ end of tRNA,making it an “activated” amino acid • Usually at least 20 per species; sometimes only 18 (no gln,asn), sometimes > 20 • Typically a synthetase can recognize many isoacceptor tRNAs Translation; Mol Bio Methods
Activation reaction • Amino acid + tRNA + ATP aminoacyl-tRNA + AMP + PPi • Attachment is to 3’ hydroxyl ofribose at the end of the terminaladenylate on the acceptor stem • Some of them actually attach at 2’ and a subsequent rearrangement moves the amino acid to the 3’ position • As usual, the reaction is driven toward completion by hydrolysis of pyrophosphate Translation; Mol Bio Methods
Energy and Recognition Each aminoacyl-tRNA synthetase has to bind ATP, for obvious reasons Aminoacyl-tRNA synthetases have to recognize the anticodon loop even though it’s at the opposite end of the tRNA molecule Requires an intimate interaction between the tRNA and the synthetase Translation; Mol Bio Methods
Ribosomes Sketch courtesy Carleton College • Protein synthesis requires: • Ribosome (catalysis) • Accessory proteins (scaffold) • mRNA (information) • Aminoacyl-tRNA (activated aa) • Every ribosome contains a small subunit and a large subunit • E.coli: 70S made up of 30S and 50S • Eukaryotes: 80S made up of 38S and 60S Translation; Mol Bio Methods
E.coli ribosomes Sketch courtesyU. Colorado • 30S subunit: • RNA 16S, 1542 nucleotides;extensive secondary structure • 21 proteins • 50S subunit: • 5S rRNA, 120 nucleotides • 23S rRNA, 2904 nucleotides • 31-34 proteins Translation; Mol Bio Methods
Structures of prokaryotic ribosomes H.marismortui 50S ribosomal subunit1405kDa oligomerPDB 1FFK, 2.4Å Thermus 30Sribosomal subunit791kDa oligomerPDB 3T1Y, 2.8Å Translation; Mol Bio Methods
Three tRNA binding sites All 3 are on the 50S subunit of ribosome P site: binds tRNA carrying existing chain A site: binds incoming aminoacyl-tRNA that is about to be added to chain E site: holds empty tRNA prior to release Translation; Mol Bio Methods
Initiation • Initiation factors (proteins) required to set up the start of synthesis • IF-1,2,3 in prokaryotes; ~8 in eukaryotes • IF-3 maintains ribosomal subunits in their dissociated state at this stage • IF-2 selects the right initatior tRNA • Once the 30S complex forms at initiation codon, 30S and 50S can come together Translation; Mol Bio Methods
Shine-Dalgarno sequences Purine-rich sequence on mRNA ~10 bases upstream of initiation AUG Binds to pyrimidine-rich sequence on 16S rRNA part of 30S subunit of ribosome Aligns that for proper translation Typical sequence: GGAGGU Translation; Mol Bio Methods
Elongation • Each activated amino acid has to be delivered to the catalytic adenine in the ribosomal RNA • Peptidyl-tRNA occupies P site • Elongation factor (EF-Tu) provides GTP binding site at “A” site • Framework near that base (RNA, protein) holds tRNA appropriately so it can deliver the amino acid to peptidyl transferase activity of rRNA • Reaction:tRNA-aa + Proteinn + GTP tRNA + GDP + Pi + Proteinn+1catalyzed and completed Translation; Mol Bio Methods
Translocation After peptide bond formed: newly created peptidyl-tRNA is partially in A site, partially in P site Empty tRNA moves somewhat out of P site into exit(E) site These have to be released and peptidyl-tRNA must move from A to P mRNA must shift by one codon relative to ribosome Translation; Mol Bio Methods
Termination Release factors (RF-1,2,3) participate in termination of synthesis at UGA,UAG, UAA No tRNAs recognizes these so synthesis stalls; release factor diffuses into A site Protein hydrolyzed off ribosome (+GTP) Ready for another round Potential for post-translational modifications Controversy remains about how quickly proteins fold up as they emerge Translation; Mol Bio Methods
iClicker question 2 2. The coding region of a gene is 1200 bp. 60% of it is introns. The number of bases in the mature mRNA transcript of the gene is • (a) 1200 bases • (b) 3600 bases • (c) 160 bases • (d) 480 bases • (e) 720 bases Translation; Mol Bio Methods
iClicker question 3 3. For the DNA just mentioned (1200 bp, 60% introns) how many amino acids will appear in the translated protein? • (a) 1200 aa’s • (b) 1440 aa’s • (c) 160 aa’s • (d) 240 aa’s • (e) 480 aa’s Translation; Mol Bio Methods
Synthesizing nucleic acid polymers • Laboratory synthesis of nucleic acids requires complex strategies • Functional groups on the monomeric units are reactive and must be blocked • Correct phosphodiester linkages must be made • Recovery at each step must high! Translation; Mol Bio Methods
Solid Phase Oligonucleotide Synthesis • Well-defined process involving attaching blocking groups to reactive components, adding extensions, unblocking, and then cycling through the various steps. Translation; Mol Bio Methods
High-throughput sequencing Some methods are large-scale automated versions of what we’ve described (Next-Gen Sequencing) (Illumina) – accurate, fast Others are based on passage of DNA through pores (3rd Gen) (Nanopore) – less accurate, even faster Translation; Mol Bio Methods
Cloning • Cloning is the process whereby DNA is copied in a controlled way to produce desired genetic results Translation; Mol Bio Methods
Plasmids • Small (typically < 10 kbp), usually circular segments of DNA that get replicated along with the organism’s chromosome(s) • Bacterial plasmids have a defined origin of replication and segments defining specific genes • Some are natural; others are man-made Translation; Mol Bio Methods
How they’re used • Typical man-made plasmid includes a gene that codes for an enzyme that renders the bacterium resistant to a specific antibiotic, along with whatever other genetic materials the experimenter or clinician wishes to incorporate • Cells that replicated the plasmid will be antibiotic-resistant; surviving colonies will be guaranteed (?) to contain the desired plasmid in all its glory Translation; Mol Bio Methods
A typical plasmid Translation; Mol Bio Methods
Building useful plasmids • Take starting plasmid and cleave it with a restriction enzyme at a specific site • Add foreign DNA that has been tailored to fit into that plasmid Translation; Mol Bio Methods
Inserts • Typically a place within the plasmid will be set up so that small stretches (< 10 kbp) of desired DNA can be ligated in • With sticky ends: high specificity, but you do get self-annealing of the plasmid and of the insert, so those have to be eliminated • With blunt ends: require more artisanry:T4 phage ligase can rejoin ends without stickiness; but it’s chaotic Translation; Mol Bio Methods
Directional cloning • Guarantees that the desired DNA goes in in exactly one orientation Translation; Mol Bio Methods