Download

1 / 59
590 likes | 630 Vues
Dive into the essential process of electron transport and oxidative phosphorylation, understanding the generation of ATP from reducing equivalents. Explore oxidation-reduction reactions, proton pumping, ATP synthesis, and the vital role of electron transport in energy production. Learn about TCA Cycle, aerobes vs. anaerobes, chemiosmotic theory, oxidation states, and the biochemical significance of energy conversions. Gain insights into the mechanisms driving ATP synthesis, cellular energy balance, and the fascinating biochemistry behind these crucial metabolic pathways.

E N D
Electron Transport and Oxidative Phosphorylation Andy HowardBiochemistry Lectures 1 November 2010 OxPhos + Electron Transport
Electron Transport • This shows how we can really make ATP from all those reducing equivalents that we amassed during glycolysis and the TCA cycle OxPhos + Electron Transport
Oxidative Phosphorylation Generalizations about oxidation-reduction reactions Electron Transport: Complexes I-IV The proton pump Driving protons across the membrane ATP synthase Bookkeeping Control What we’ll discuss OxPhos + Electron Transport
iClicker quiz, question 1 • Why does it matter that the G6PDH gene is located on the X chromosome? (a) males don’t possess the gene (b) females don’t possess the gene (c) only one copy available per cell (d) no DNA-repair mechanisms available for X-Chromosome genes OxPhos + Electron Transport
iClicker question 2 A reaction that replenishes an intermediate is said to be • (a) anabolic • (b) catabolic • (c) anapleurotic • (d) amphibolic • (e) none of the above. OxPhos + Electron Transport
TCA Cycle and Evolution • The entire pathway didn’t evolve together • Some reactions much older than others • Some ran backward in early implementations • Several enzymes adapted from amino acid degradation • Youngest enzyme:-ketoglutarate dehydrogenase OxPhos + Electron Transport
Aerobes and anaerobes • Because of close coupling between TCA cycle and oxidative phosphorylation, the complete TCA cycle is an aerobic phenomenon • Anaerobes do have most of these enzymes, but the sequence of reactions is different • Oxygen is actually toxic to many anaerobes OxPhos + Electron Transport
Overall role of electron transport • Last 3 lectures: we discussed carbohydrate metabolism and the TCA cycle, each of which produced some high-energy phosphate energy directly. • In both of those systems much of the energy generated took the form of reduced cofactors--NADH in both systems, and FADH2 (or QH) in the Krebs cycle. • Now we’ll see what happens to those! OxPhos + Electron Transport
The overall reactions • NADH + H+ + (1/2)O2 + 2.5 ADP+ 2.5 Pi NAD+ + H2O + 2.5 ATP • ETS also catalyzes transformations of the flavin coenzyme FAD:FADH2 + (1/2)O2 + 1.5 ADP + 1.5 Pi FAD + H2O + 1.5 ATP • These are mediated through other cofactors: Q, cytochromes, and Fe-S proteins • Proton translocation is crucial OxPhos + Electron Transport
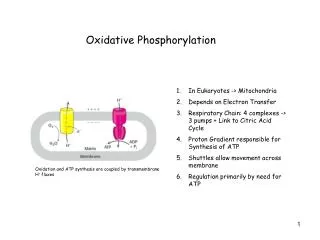
Chemiosmotic theory:What it says • Protons are translocated from outside of mitochondrial inner membrane into its interior • That passage actually generates both chemical and electrical energy. • This is because they are moving down a concentration and electrical-potential gradient:Remember:Gtrans = RTln[Cin]/[Cout] + zF OxPhos + Electron Transport
How it works • This energy is used to drive the synthesis of ATP from ADP and Pi within an enzyme called ATP synthase, which is (big surprise!) anchored on the inside of the inner mitochondrial membrane. • The structure of two components of this enzyme system were determined in 1999 by Andrew Leslie and others. OxPhos + Electron Transport
Oxidation state and energy • We typically measure oxidation states in volts. • We can relate the energy associated with an oxidation-reduction reaction—the so-called change in redox potential—with the change in the oxidation state of the molecules involved in the reaction. OxPhos + Electron Transport
What is a volt? • A volt is actually a measure of energy per unit charge; in fact, a volt is one joule per coulomb. • When we say that a double-A battery has a voltage of 1.5 V, we mean that it can (under optimal conditions) deliver 1.5 joules of energy( = 0.359 cal, or 3.59*10-4 kcal) per coulomb of charge. OxPhos + Electron Transport
Charge and energy • One electron carries a charge of1.602 * 10 -19 coulomb • If change in redox potential in a reaction is 0.32 V and all of that change is delivered to a single electron:then energy imparted to that electron is • eΔE =(1.602 * 10-19 coulomb / e-) *(0.32 J/coulomb)= 0.513*10-19J / e- = 1.23* 10 -23 kcal / e- OxPhos + Electron Transport
… in biochemical units … • That doesn't sound like much, but if we look at that on a per mole basis, 0.32V applied to a single electron is(1.23 * 10-23 kcal/e-) *6.022 * 1023 e -/mole= 30.87 kJ/mol = 7.38 kcal/mol • which is a reasonable amount of energy on the scale we're accustomed to examining (~1 ATP hydrolysis). OxPhos + Electron Transport
So what can we get? • There is enough energy bound up in the reduced state of NAD relative to the oxidized state to drive the net creation of 2.5 molecules of ATP from ADP and phosphate, as indicated in the equations shown above. • Since there are NADH molecules created in several steps in glycolysis and the Krebs cycle, there numerous net ATP molecules that arise from the overall process. OxPhos + Electron Transport
Results from TCA cycle • 3 NADH produce 7.5 ATP • 1 FADH2 produces 1.5 ATP • 1 substrate-level phosphorylation(from succinyl CoA hydrolysis) • Total: 10 ATP per round, if we don’t get interrupted! • Since we formally shove 2 molecules of acetyl CoA into the system per molecule of glucose input into glycolysis, we get 20 ATPs out of the TCA cycle per glucose. OxPhos + Electron Transport
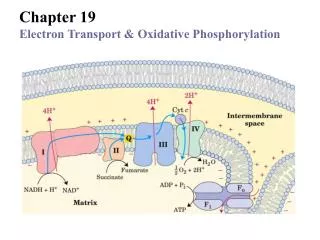
ETS: The big picture • 5 membrane-associated, multi-enzyme complexes in mitochondrial inner membrane • Complexes I-IV associated with electron transport and proton translocation:Complexes I, III, IV move protons • Complex V uses that proton gradient to produce ATP from ADP and Pi OxPhos + Electron Transport
Complexes I-IV • There are several multi-enzyme complexes involved in converting the reductive energy in NADH to its final products. # Name I NADH-Ubiquinone oxidoreductase II Succinate-ubiquinone oxidoreductase III Ubiquinol-cytochrome c oxidoreductase IV Cytochrome c oxidase OxPhos + Electron Transport
Overview of Oxidative Steps Chart courtesyMichael King,Indiana State OxPhos + Electron Transport
Reduced cofactors to ATP • We will discuss how the energy latent in these reduced cofactors can be turned into energy in the form of high-energy phosphate bonds in nucleoside triphosphates--the standard currency of energy. OxPhos + Electron Transport
What the ETS does • The electron transport system (ETS) is responsible for these transformations. • Like the Krebs cycle or glycolysis, the electron transport chain is a series of chemical transformations facilitated by proteins. OxPhos + Electron Transport
Roles of these systems • Some of these proteins are enzymes in the conventional sense • others are not—they're electron transport proteins only: • so they can only be regarded as enzymes if we allow that the entire ETS is a large, multi-polypeptide transformation system--a multi-component enzyme, analogous to the pyruvate decarboxylase complex or the fatty acid synthase complex OxPhos + Electron Transport
Flavin cofactors • Participants in ETS • Sometimes depicted as starting points, but it’s probably better to think of them as intermediaries OxPhos + Electron Transport
Complex I • NADH:Ubiquinone oxidoreductase • Embedded in inner mitochondrial membrane • Passes electrons from NADH to ubiquinone: OxPhos + Electron Transport
Properties of ubiquinone • The quinone moiety is where the oxidation-reduction takes place(1e- or 2e- at a time) • The hydrophobic tail (15C = 3 isoprene units) anchors this cofactor to the mitochondrial membrane OxPhos + Electron Transport
Protons in Complex I • Complex I picks up a pair of protons from the matrix and passes them up through to the other side of the membrane • Energy for this translocation (against the concentration gradient) supplied by oxidation of NADH Diagram courtesy of Rice University OxPhos + Electron Transport
Complex II • Succinate-ubiquinone oxidoreductase • We’ve looked at this already as succinate dehydrogenase • Succinate + Q Fumarate + QH2 • No protons translocated in this step:Reaction is close to isoergic • FAD and Fe-S proteins involved PDB 2FBW246 kDadimer of hetero-tetamersChicken OxPhos + Electron Transport
Complex III • Ubiquinol:cytochrome c oxidoreductase • Transfers electrons from reduced ubiquinol to iron atoms in cytochrome c • One electron per cytochrome c • Enzyme contains three main subunits: cytochrome c1, cytochrome b,and an iron-sulfur (“Rieske”) protein • Net reactions: • QH2 + cyto c–Fe3+ QH• + cyto c–Fe2++ H+ • QH• + cyto c–Fe3+ Q + cyto c–Fe2+ + H+ OxPhos + Electron Transport
Complex III structures • Cytochrome bc1 complex • Rieske Fe-S protein OxPhos + Electron Transport
Cytochrome-dependent steps • Cytochromes are, in general, proteins involved in electron transport. • Cytochrome c is a mobile, soluble compound • Others are generally membrane-associated • What they have in common:covalently bound heme OxPhos + Electron Transport
Cytochromes • The name derives from the fact that many of them are colored (–chrome)and found in cells (cyto–). • In particular, cytochrome c, which is a significant intermediary in the ETS, is a water-soluble, relatively small, heme-containing protein • Cytochrome c received substantial attention in the early years of biochemistry both because of its inherent importance and because it's easy to study. OxPhos + Electron Transport
Where do the individual names come from? • Early analyses of cellular or mitochondrial extracts showed several peaks in the absorption spectra • Cytochromes were named as those spectral peaks became associated with specific spectral peaks • Sometimes two different species (e.g. cytochrome c and cytochrome c1) have very similar spectra OxPhos + Electron Transport
Cytochrome c and evolution Cytochrome c is highly conserved • rate of mutation across the billions of years of evolution is remarkably slow, as compared to other proteins. • This is generally a sign that its function is sufficiently irreplacable that even a modest modification in the protein renders the cell unviable. OxPhos + Electron Transport
Cytochrome c structure • Monomer is only 12kDa • Heme group appears between helices • Covalent linkage to heme visible here • Highly soluble aqueous protein, unlike the larger, membrane bound cytochromes Yeast cytochrome cPDB 1YCC, 12.8 kDa1.23Å; dimer shown OxPhos + Electron Transport
Cytochrome c and apoptosis • Cytochrome c plays a role in apoptosis: • When a cell receives (and pays attention to) an external apoptotic signal, typically via IP3 signaling pathway, Ca2+ is released from the ER • One of the first responses is release of cytochrome c from the mitochondrion into the cytosol • This triggers lytic events that eventually lead to shrinkage and absorption of the cell fragment OxPhos + Electron Transport
Complex 3: schematic CourtesyU.Texas OxPhos + Electron Transport
Complex IV • Cytochrome c oxidase • Transfers electrons from (soluble) cytochrome c to molecular oxygen:Product is water • 2Cyto c–Fe2+ + (1/2)O2 + 2H+ 2Cyto c–Fe3+ + H2O OxPhos + Electron Transport
Cytochrome oxidase • 2 functional units; • Up to 13 subunits containing membrane-spanning helices • 4 protons produced per oxidation of two molecules of QH2 at the Q0 site • See fig. 14.13 OxPhos + Electron Transport
Cytochrome oxidase mechanism • Depends on two Cu+ ions www.steve.gb.com OxPhos + Electron Transport
Mitochondrial sequestration • Mitochondrion is a fairly complex organelle, found in all eukaryotes. • Some simple algae have one mitochondrion per cell, whereas some protozoa have a half-million per cell. • A mammalian liver cell contains about 5000 mitochondria. These organelles resemble bacteria in size and complexity. OxPhos + Electron Transport
Mitochondrial genetics • Vertebrate mitochondrion has its own chromosome, but it does not code for many proteins • Human mitochondrion codes for 17 proteins, plus two dozen specialized tRNAs and (presumably) some control elements. OxPhos + Electron Transport
Where is the rest of the mitochondrion’s function specified? • Those 17 proteins are ~2% of the functioning proteins in the mitochondrion • Most of the ~1000 proteins that function in the mitochondrion are coded for in the host genome and are translocated, sometimes with some amount of proteolytic processing, from the ribosomal protein-synthesis mechanisms of the cytosol into the interior of the mitochondrion. OxPhos + Electron Transport
Mitochondrial structure • The mitochondrion has asmooth outer membraneand a highly convoluted innermembrane • Intermembrane space between them • Outer membrane is permeable to small molecules, so functionally the intermembrane space is equivalent to the cytosol. OxPhos + Electron Transport
Mitochondrial localization • In eukaryotes, ETS and Krebs reactions take place in the mitochondrion. • Many reactions occur on the inner membrane in its folded surfaces called cristae. • Localization to the membrane provides for orderly passage of substrates or electrons from one protein to the next, helping to defeat old man entropy. OxPhos + Electron Transport
Mitochondrial matrix enzymes Some proteins do function in the matrix, the aqueous compartment of the mitochondrion interior to the inner membrane. • Pyruvate dehydrogenase complex • TCA-cycle enzymes (except succinate dehydrogenase, which is embedded in the inner membrane) • Some enzymes involved in fatty acid oxidation. OxPhos + Electron Transport
What’s the matrix like? • This matrix is has such a high overall protein concentration that it is not really an aqueous medium; it's a gel. • Think of reactions that occur in the mitochondrial matrix as occuring in Karo syrup (except the syrup is made of protein + H2O, not sugar + H2O) OxPhos + Electron Transport
Regulation of ETS • An element of regulation of electron transport is based on the availability of adenosine diphosphate, the substrate for the oxygen-mediated respiration steps. Mitochondria consume oxygen rapidly when ADP is available. OxPhos + Electron Transport
Uncouplers • When all the ADP in a mitochondrion is consumed, rapid respiration stops. • There are compounds that uncouple oxidation from phosphorylation • when these uncouplers are added to a test system, it will continue to oxidize NADH even in the absence of ADP. • 2,4-dinitrophenol is an example of an uncoupler. OxPhos + Electron Transport
ATP synthase • Crucial example of a molecular motor, i.e.,a machine that translates between chemical energy and mechanical work • Ultimately its job is to pull protons across the membrane, using the energy associated with that favorable translocation to drive the synthesis of ATP from ADP and Pi • The motor is what rotates the protein through various positions to enable its reactions OxPhos + Electron Transport