Download

1 / 30
370 likes | 518 Vues
This chapter discusses the intricate process of oxidative phosphorylation, a key stage in cellular respiration where energy from reduced fuels like carbohydrates, lipids, and amino acids is harnessed to synthesize ATP in animals. It highlights the role of NADH and FADH2 in transferring electrons through the electron transport chain, leading to ATP production. The chemiosmotic theory is explained, underscoring the importance of proton gradients across mitochondrial membranes. The chapter also details the structure of mitochondria and the fundamental components of the electron transport chain.

E N D
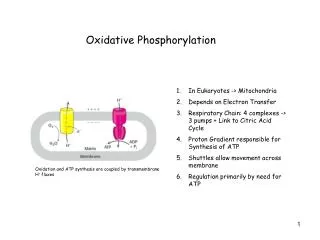
Oxidative Phosphorylation Chapter 19 (Page 690-718)
1. Energy from Reduced Fuels is used to Synthesize ATP in Animals • Carbohydrates, lipids, and amino acids are the main reduced fuels for the cell. • Electrons from reduced fuels are transferred to reduced cofactors NADH or FADH2. • In oxidative phosphorylation, energy from NADH and FADH2 are used to make ATP.
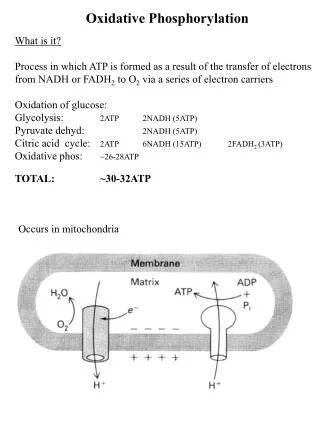
2. Energy Flow in Cellular Respiration • Stage 1: Oxidation of metabolic fuels yielding acetyl-CoA. • Stage 2: Oxidation of acetyl groups in the citric acid cycle includes four steps in which electrons are abstracted. • Stage 3: Electrons carried by NADH and FADH2 are funneled into a chain of mitochondrial electron carriers-the respiratory chain-ultimately reducing O2 to H2O. • e- flow drives the production of ATP
3. Oxidative Phosphorylation • Electrons from the reduced cofactors NADH and FADH2 are passed to proteins in the respiratory chain. • In eukaryotes, oxygen is the ultimate electron acceptor for these electrons. • Energy of oxidation is used to phosphorylate ADP generating ATP. • This process was discovered in 1948 by Eugene Kennedy and Albert Lehninger.
3A. Chemiosmotic Theory ADP + Pi ATP Highly Thermodynamically Unfavorable How do we make it possible? • Phosphorylation of ADP is not a result of a direct reaction between ADP and some high-energy phosphate carrier. Rather the process involves: • Energy needed to phosphorylate ADP is provided by the flow of protons down the electrochemical gradient • The energy released by electron transport is used to transport protons against the electrochemical gradient
3A. Chemiosmotic Theory Four e- carrying complexes. ATP Synthase Complex
3B. Chemiosmotic Energy Coupling Requires Membranes • The proton gradient needed for ATP synthesis can be stably established across a membrane that is impermeable to ions. • Inner membrane in mitochondria • Membrane must contain proteins that couple the energetically “downhill” flow of electrons in the electron-transfer chain with the energetically “uphill” flow of protons across the membrane. • Membrane must contain a protein that couples the “downhill” flow of protons to the phosphorylation of ADP.
4. Structure of a Mitochondrion Double membrane leads to four distinct compartments: • Outer Membrane: • Relatively porous membrane allows passage of metabolites • Intermembrane Space (IMS): • similar environment to cytosol • higher proton concentration (lower pH) • Inner Membrane • Relatively impermeable, with proton gradient across it • Location of electron transport chain complexes • Convolutions called Cristae serve to increase the surface area • Matrix • Location of the citric acid cycle and parts of lipid and amino acid metabolism • Lower proton concentration (higher pH)
5. Electron-transport Chain Complexes contain a Series of Electron Carriers • Each of the 4 complexes contains multiple redox centers consisting of: • Flavin Mononucleotide (FMN) or Flavin Adenine Dinucleotide (FAD) • Initial electron acceptors for Complex I and Complex II • Can carry two electrons by transferring one at a time • Tightly bound, sometimes covalently, to flavoproteins • Standard reduction potential is dependent on protein environment • Cytochromes a, b or c • Iron-sulfur cluster
5A. Cytochromes • One electron carriers (Fe2+ ↔ Fe3+) • Iron coordinating porphoryin ring derivatives • a, b or c are integral proteins but differ by ring additions • c also has peripheral protein form
5 B. Iron-Sulfur Clusters • One electron carriers • Coordinate Fe2+ by cysteines in the protein and inorganic S atoms • Centers defined by inorganic S coordination • Contain equal number of iron and sulfur atoms • Standard reduction potential of the iron varies depending on the center and interaction with protein. 1 Fe 2Fe-2S 4Fe-4S
6. Ubiquinone (Coenzyme Q) • Ubiquinone is a lipid-soluble conjugated dicarbonyl compound that readily accepts electrons • Upon accepting two electrons, it picks up two protons to give an alcohol, ubiquinol • Ubiquinol can freely diffuse in the membrane, carrying electrons with protons from one side of the membrane to another side • Coenzyme Q is a mobile electron carrier transporting electrons from Complexes I and II to Complex III
7. Free Energy of Electron Transport Reduction Potential (E) ∆Eo′ = Eo′(e- acceptor) – Eo′(e- donor) ∆Go′ = –nF∆Eo′ For negative G need positive E E(acceptor) > E(donor) Electrons are transferred from lower (more negative) to higher (more positive) reduction potential. Free Energy released is used to pump protons, storing this energy as the electrochemical gradient.
7. Free Energy of Electron Transport Order of e- can be deduced from E’°: NADH → Cyt a3 → Q → Cyt b → Cyt c1 → Cyt c → Cyt a → O2
8A. Flow of Electrons from Biological Fuels into the Electron-Transport Chain
8B. NADH:ubiquinone oxidoreductase, a.k.a. Complex I • L-shaped complex and starting point for NADH for electron transfer. • One of the largest macro-molecular assemblies in the mammalian cell. • Over 40 different polypeptide chains, encoded by both nuclear and mitochondrial genes.
8B. NADH:ubiquinone oxidoreductase, a.k.a. Complex I • NADH binding site located in the matrix side. • Noncovalently bound flavin mononucleotide (FMN) accepts two electrons from NADH • Several iron-sulfur centers pass one electron at a time toward the ubiquinone binding site
8BI. Complex I- NADH to Ubiquinone NADH NAD+ + H+
8BII. NADH:Ubiquinone oxidoreducase is a proton pump • Transfer of two electrons from NADH to ubiquinone is accompanied by a transfer of protons from the matrix (N for loss of protons) to the intermembrane space (P for gain of protons) • Experiments suggest that about four protons are transported per one NADH NADH + Q + 5H+N → NAD+ + QH2 + 4 H+P • Reduced coenzyme Q picks up two protons
8C. Succinate Dehydrogenase, a.k.a. Complex II • Complex II is the starting point for FADH2 for electron transfer. • FAD accepts two electrons from succinate (recall the citric acid cycle).
8C. Succinate Dehydrogenase, a.k.a. Complex II • Electrons are passed, one at a time, via iron-sulfur centers to ubiquinone, which becomes reduced QH2. • Does not transport protons.
8CI. Complex II- Succinate to Ubiquinone • The complex consists of 4 subunits, two of which (C and D) are transmembrane. • Heme b is not on the main path of e- transfer but may protect against reactive oxygen species.
8D. Ubiquinone:Cytochrome c Oxidoreductase, a.k.a. Complex III • Uses two electrons from QH2 to reduce two molecules of cytochrome c. • Involves iron-sulfur clusters, cytochrome bs, and cytochrome cs. • Engages in a Q cycle, which results in four additional protons being transported to the intermembrane space.
8DI. Complex III- Ubiquinone to Cyt C • The complex is a dimer of identical monomers, each with 11 different subunits. • The complex has two distinct binding sites for ubiquinone, QN and Qp. • Rieske iron-sulfur proteins have Fe-S clusters with His residue attachments in place of Cys.