Download

1 / 53
530 likes | 721 Vues
Serial Regulation of Transcriptional Regulators in the Yeast Cell Cycle Cell, Vol 106, 697-708, September 2001 E2F integrates cell cycle progression with DNA repair, replication, and G2/M checkpoints Genes & Development, Vol 16, 245-256, 2002. ניתוח רשתות רגולטוריות - הקדמה.

E N D
Serial Regulation of Transcriptional Regulators in the Yeast Cell Cycle Cell, Vol 106, 697-708, September 2001 E2F integrates cell cycle progression with DNA repair, replication, and G2/M checkpoints Genes & Development, Vol 16, 245-256, 2002
ניתוח רשתות רגולטוריות - הקדמה שני המאמרים עוסקים ביצירת תמונה ברורה ושלמה יותר של הרשת הרגולטורית במחזור התא – בשמר ובאדם. שניהם עושים זאת בעזרת שיטה שפותחה בכדי להפוך הישג זה לאפשרי.
המגבלות של הכלים המחקריים קיימות שתי שיטות לזיהוי השפעה של פקטור שעתוק על גן מסויים: (1) ניתוח השינוי בביטוי הגן כתוצאה מהוספת פקטור שעתוק מסויים (microarray analysis). הבעיה – האם זו השפעה ישירה של הפקטור ? (2) מציאת אתרי קישור של הפקטור לגן מסויים. הבעיה - האם הפקטור אכן מבקר את הגן ?
הפתרון: Genome-Wide Location Analysis השיטה משלבת בין מציאת אתרי קישור ישירים בין הפקטורים לדנ"א תוך שימוש ב- IP (chromatin immunoprecipitation), לביןרמת ביטוי חלבונים (ע"י DNA microarray analysis). שילוב השיטות מנטרל את החסרונות של שתיהן, ומאפשר בכך ניטור מדוייק יותר של אינטרקציות בין דנ"א לחלבונים.
תיאור השיטה קישור החלבונים לדנ"א בנוכחות formaldehyde ("קיבוע" הקישור בין החלבונים לדנ"א). שבירת התאים ע"י סוניקציה לצורך הפקת הדנ"א.
תיאור השיטה (המשך) • הפרדת מקטעי הדנ"א הרלוונטים (שנקשר אליהם החלבון הנחקר) מכלל הדנ"א התאי ע"י Immunoprecipitation (העברת הדנ"א דרך קולונה עם נוגדנים ספציפיים לחלבון).
דנ"א ביקורת דנ"א שעבר IP תיאור השיטה (המשך) • הפרדת הנוגדנים והחלבונים מהדנ"א. • אמפליפיקציה וצביעה פלורוסנטית של מקטעי הדנ"א שהופרדו ב – Cy5 (אדום), ע"י LM-PCR. • אמפליפיקציה וצביעה פלורוסנטית של מקטעי דנ"א מדגימת ביקורת (שלא עברה IP) ב – Cy3 (ירוק), ע"י LM-PCR.
תיאור השיטה (המשך) • היברידיזציה של מקטעי הדנ"א המסומנים בשני הצבעים ל- microarray. microarray
עיבוד תוצאות ההיברידיזציה הניסוי בוצע שלוש פעמים ובסופו של דבר התוצאות שוקללו כדי לחשב את מידת הקישור היחסית של החלבון לכל אחד מהרצפים ב- microarray. צביעה באדום משמעותה שמקטע הדנ"א הספציפי על ה- microarray מכיל אתר קישור לחלבון.
עיבוד תוצאות ההיברידיזציה (המשך) כל נקודה בגרף מייצגת מקטע ב- microarray והגרף משווה בין עוצמת הצביעה שלו באדום לבין זו בירוק. המקטעים שיצוגם חורג מגבולות ה- p-value הם מקטעים שעוצמת צביעתם באדום גדולה יותר (ההסתברות לקישור מקרי קטנה מ- 0.001).
Serial Regulation of Transcriptional Regulators in the Yeast Cell Cycle הקדמה: עד כה זוהו 9 פקטורי שעתוק בשמר הקשורים לבקרת גנים במחזור התא, אולם אין מידע מלא על קבוצת הגנים שכל אחד מהם מבקר. מי מבקר את המבקרים? כיצד עובדת הבקרה על פקטורי השעתוק עצמם? רגולציה מתואמת על גנים בעלי פונקציונליות משותפת וספציפית לשלב מסויים.
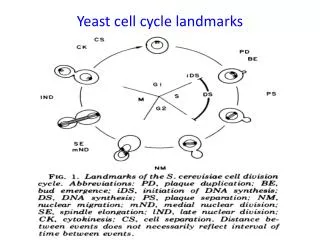
מחזור התא בשמר היפרדות הכרומוזומים לשני תאי הבת וציטוקיניזה שלב המפריד בין סוף המיטוזה לבין התחלת הכפלת הדנ"א G0 הכפלת הדנ"א והתחלת ההנצה מפריד בין סוף ההכפלה לתחילת המיטוזה
מהלך הניסוי שימוש בשיטת genome-wide location analysis כפי שתוארה קודם, על מנת לזהות את אתרי הקישור in vivo של כל אחד מתשעת פקטורי השעתוק. מתוך כלל הגנום של השמר, בודדו כ- 800 גנים שרמת הביטוי שלהם משתנה באופן מחזורי עם התא. אתרי הקישור שזוהו סווגו ע"י p-value שונים, כאשר ההתמקדות היתה ב- 213 אתרים בעלי p<0.001 (מהווים 27% מ- 800 הגנים הנ"ל). אימות התוצאות ע"י בדיקת רמות ביטוי.
מהלך הניסוי (המשך) בלי IP (ביקורת) עם IP
SBF MBF תוצאות • שילוב פקטורי השעתוק בבקרה של מחזור התא: • לפני מחקר זה נבנה מודל כללי, על סמך מספר קטן יחסית של גנים. המחקר אישש מודל זה, והרחיב אותו. המודל הכללי שהיה קיים
ידע שנוסף ידע קודם בקרה על פקטורי השעתוק תוצאות הניסוי איששו ואף הרחיבו את המידע לגבי מעורבותם של פקטורי שעתוק בבקרה על פקטורים בשלב הבא במחזור התא. התוצאות חשפו תמונה של מעגל רגולטורי שלם שיוצרים הפקטורים זה עם זה.
רגולציה על הגנים של Cyclin/CDK מעבר בין שלבים במחזור התא תלוי בשינויים בפעילותם של קומפלקסי Cdc28-cyclin. הגילוי: פקטורי שעתוק המבקרים ציקלינים בשלב מסויים, מבקרים גם גנים המקודדים לרכיבי מפתח המעורבים במעבר לשלב הבא במחזור התא. מידע חדש זה מסייע בהבנת תרומתה של בקרת השעתוק להתקדמותו של מחזור התא.
רגולציה של פונקציות ספציפיות לשלב • תוצאות הניסוי סייעו בהגדרת קבוצות גנים, שכל אחת מהן אחראית על פונקציה ספציפית במחזור התא, והתאימה לכל קבוצה כזו את פקטורי השעתוק שלה.
חפיפת בקרה בין פקטורי השעתוק תאי שמר עם מוטציה ב- Mcm1 או ב- Ndd1 אינם חיוניים, אולם מוטציה באחד משבעת הפקטורים האחרים אינה פוגעת בחיוניות התא. ההסבר ההגיוני - חפיפה בין קבוצות הגנים המבוקרות ע"י זוגות פקטורים שונים. מחקר זה הראה כי חפיפה זו אינה נוצרת כתוצאה מהמוטציה בלבד, אלא כי חפיפה חלקית קיימת גם בתאים נורמליים. החפיפה החלקית מסבירה כיצד פקטור אחד יכול לחפות על דפקט בבן זוגו, אך עדיין לשמור על פונקציונליות יחודית ונבדלת מזו של בן הזוג.
חפיפת בקרה בין פקטורי השעתוק (המשך) • חפיפה בין אתרי המטרה של זוגות פקטורים הומולוגים. • חפיפה בין אתרי המטרה של זוגות פקטורים המשתייכים לאותו הקומפלקס. • חפיפה בין אתרי המטרה של זוגות פקטורים שהקשר ביניהם לא היה ידוע.
דיון בתוצאות המאמר • ע"י שילוב של genome-wide location analysis ו- expression analysis זוהו המטרות הגנומיות של כל 9 פקטורי השעתוק הידועים במחזור התא בשמר. התוצאות הראו שרשת רגולטורית מקושרת ומעגלית מבקרת את מחזור התא ברמת השעתוק.
דיון בתוצאות המאמר (המשך) • נקודות עיקריות שעלו במחקר : • רשת הבקרה על פקטורי השעתוק עצמם הינה מעגלית – תופעה זו חשובה מכיוון שהיא מעניקה הבנה נוספת על האופן בו התאים מבטיחים מעבר משלב לשלב, ומכיוון שהיא מהווה פתח למחשבה שתופעות כדוגמתה קיימות גם ביצורים מפותחים יותר.
דיון בתוצאות המאמר (המשך) • נקודות עיקריות שעלו במחקר (המשך) : • חלק מהפקטורים מבקרים הן על כניסה לשלב מסויים (ע"י בקרה על ה- cyclins שלו), והן על היציאה ממנו (ע"י בקרה על המעבר לשלב הבא). • תמונה שלמה יותר של מחזור התא מתקבלת ע"י שילוב של מידע שהיה ידוע קודם לכן, עם מידע שהתקבל במאמר.
דיון בתוצאות המאמר (המשך) • חשיבות המידע על אתרי קישור ישירים • בדיקת ביטוי הגנים לבדה אינה מספקת תמונה מדוייקת על מערכת הבקרה. • מידע על אתרי קישור מאפשר זיהוי של רצפי קונצנזוס באזורי בקרה: • המצאותו של רצף קונצנזוס בגן מסויים, אינה מעידה בהכרח על קישור הפקטור לשם, לכן לצורך הוכחה חותכת יש צורך במידע על אתר קישור ישיר. • יתכן מצב שבו פקטור נקשר לאתר שאינו מכיל את רצף הקונצנזוס שזוהה עבורו - מידע על אתרי קישור ישירים מספק הסבר לכך.
תרומה עתידית של המחקר שילוב מידע על קישור ישיר וביטוי כפי שהוצג במאמר מהווה פתח לגישות חישוביות חדשות שמטרתן זיהוי רשתות בקרה גנטיות. לדוגמה: • שימוש במידע על קישור להצעת זוגות פקטור-אתר מטרה עם p-values שונים, שיציבו מגבלות על המודלים האפשריים לרשתות בקרה. • מגבלות אלו יאפשרו חיפוש מונחה ולכן יעיל יותר של רשתות רגולטוריות על בסיס רמת ביטוי גנים.
בתיאבון קהל נכבד, באולם זה הושקעו מאמצים רבים לרווחתכם, אנא שמרו על תקינות הציוד ועל שלמותו.
E2F integrates cell cycle progression with DNA repair, replication, and G2/M checkpoints מבוא: משפחת פקטורי השעתוק E2F הינה בעלת תפקיד מרכזי במהלך מחזור התא. עד למחקר זה שיערו שהפקטורים ממשפחה זו מאקטבים גנים בשלב G1 ו- S.
מבוא (המשך) • המשפחה מורכבת מ- 6 פקטורים (E2F1 – E2F6), המחולקים לאקטיבטורים (E2F1 – E2F3) ולרפרסורים (E2F4, E2F5) – יתכן שפקטורים אלה מאקטבים שעתוק בנסיבות מסויימות. • תפקיד חשוב של E2F הוא גיוסם של חלבונים ממשפחת ה- tumor suppressorRetinoblastoma –pRB (p107 ו- p130).
Late M Late G1 Cdk2-cyclin A Rb + E2F P Active E2F Genome Enzymes for DNA synthesis G1 S Inactive complex
חלבונים אלה נקשרים ל- E2F ומעכבים אותם - כלומר מונעים מהם מלאקטב שעתוק של גנים בשלב G1 שיש להם תפקיד במעבר לשלב S, שבו מוכפל הדנ"א. באופן זה חלבונים אלה מונעים התפתחות סרטן. כיצד התבצע המחקר עד כה ? השיטות ששימשו עד כה לגילוי אתרי המטרהשל E2F ו- pRB מתבססות על ניתוח של ביטוי גנים. למרות ששיטות אלו השיגו התקדמות משמעותית, יש להן מספר חסרונות: קשה לקשר בודאות בין שינויים החלים בעקבות ביטוי מוגבר של הפקטורים, לבין קישורם לפרומוטורים של הגנים הרלוונטיים. ביטוי מוגבר של E2F גורם להתקדמות במחזור התא ולשינויים משניים המקשים איתור ודאי של גנים המושפעים ממנו ישירות.
הפתרון : שיטות המשלבות IP שיטות כאלה כפי שראינו במאמר הקודם, פותחו עבור מחקר דומה בשמר, אולם השיטות הללו אינן מותאמות למחקר בתאי אדם: • לא ברור האם השיטות שעבדו איתן בשמר יפיקו תוצאות נקיות מספיק מרעש בגנומים מורכבים יותר. • הגנום האנושי רוּצף, אך עדיין לא מופו כל אתרי הבקרה – לא ניתן לייצר microarray הכולל את כל אתרי הפרומוטורים בגנום.
הפתרון : שיטות המשלבות IP(המשך) • המאמר מציע גרסה מותאמת יותר של השיטה, שמאפשרת זיהוי של פרומוטורים הנקשרים ע"י פקטורי שעתוק ספציפיים בתאי אדם. • המחקר התמקד בפקטורי E2F ובכ- 1200 אתרי proximal promoters של גנים אשר ביטויים תלוי במחזור התא, מהם נבנה ה- microarray. • המחקר העלה כי ל- E2F תפקיד לא רק במעבר מ- G1 ל- S, אלא מעורב בנקודות ביקורת רבות במהלך מחזור התא.
תיאור הניסוי: • בדיקת מידת ההקשרות של E2F4 לפרומוטורים ב- G0, בכדי לאמת ממצאים קודמים לפיהם הפקטור מגייס את p130 לצורך דיכוי גנים ביציאה ממחזור התא. • הניסוי בוצע על תאים "מושתקים" (ביציאה ממחזור התא), בשיטת genome-wide location analysis דומה לזו ששימשה בשמר, עם המודיפיקציות הבאות: • על מנת להתגבר על מורכבות הדנ"א האנושי קבעו תנאים מגבילים להיברידיזציה על ה- microarray. • בניית microarray חדש שמכיל 1444 רצפי גנים (הרצפים נבחרו על פי ידע מוקדם אודות אופי אתרי הקישור של E2F)
תוצאות הניסוי 127 גנים זוהו כמטרות של E2F4 עם p-value ≤ 0.002.
אימות התוצאות גן בקרה חדשים נוגדן לא ספציפי ל- E2F4 • רבים מהגנים שהיו מוכרים כאתרי מטרה של E2F אכן זוהו בניסוי כאתרי מטרה. • אימות זיהוי הפרומוטורים כאתרי מטרה של E2F נעשה בשיטת ChIP הקונבנציונלית. • העשרה משמעותית במטרות של E2F מול העשרה זניחה ביתר הגנים.
חפיפה בין גני המטרה של E2F1 ו- E2F4 • ידע קודם מצביע על כך ש- E2F1 נקשר לגנים במעבר G1/S, ולפיכך ניסוי דומה התבצע על הפקטור בתאים בתחילת שלב S. • תוצאות הניסוי הצביעו על חפיפה משמעותית בין הגנים ששני הפקטורים נקשרו אליהם (כ- 50 גנים). • לצורך אימות התוצאות נעשתה בדיקת ביטוי הגנים על microarray שהכיל את אותם הרצפים.
גני מטרה של חדשים E2F • הניסוי חשף מספר גדול של גני מטרה שלא זוהו קודם לכן, בעלי פונקציונליות של תיקון דנ"א, נקודות ביקורת של השלבים G2 ו- M. • נעשה חיפוש של רצפי קונצנזוס לקישור פקטורי E2F. התגלה שבחלק גדול מגני המטרה קיימים מספר אתרי קישור לפקטורים, לעומת זאת בכמה מהם לא היה אף רצף כזה.
נחיצותם של p107 ו- p130 לפעילות E2F4 • בתאי עכבר עם מוטציות ב- p107 ו- p130, שעתוקם של חלק מגני המטרה של E2F4 לא דוכא. • עובדה זאת אומתה ע"י בדיקת ביטוי גנים שזוהו קודם לכן ע"י שיטת ה- genome-wide location כגני מטרה של E2F4.
+ בקרה - אימות הממצא
סקירת תוצאות המאמר • תרומת המאמר בחקר רשת הבקרה במחזור התא: • מתוצאות מחקרים קודמים המתבססים על ניתוח ביטוי גנים, התקבלו קבוצות פונקציונליות של גנים המבוקרים ע"י E2F. • אימות תוצאות קודמות – גנים העוסקים בבקרה על מחזור התא. • הרחבת תוצאות קודמות - גנים העוסקים ברפליקציה של דנ"א.
סקירת תוצאות המאמר (המשך) • בדיקת תיאוריות חדשות ואימותן – למשפחת E2F יש תפקיד גם מעבר לשלב S בניגוד למה שסברו (ויכלו להוכיח) בעבר. • הסברה שלגנים של תיקון דנ"א תפקיד חשוב בזמן ההכפלה מחוזקת ע"י מציאתם של גנים כאלה במחקר. • גילויים לא צפויים – E2F נקשר לגנים שמהווים נקודות ביקורת במחזור התא. • לדוגמא: p53 שמשמש לעצירת מחזור התא ב- G1 כתגובה לפגיעה בדנ"א.