Download

1 / 28
310 likes | 988 Vues
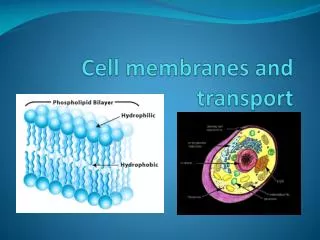
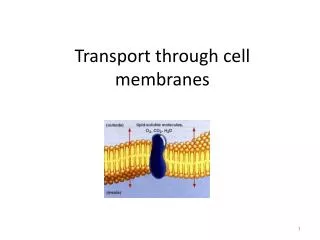
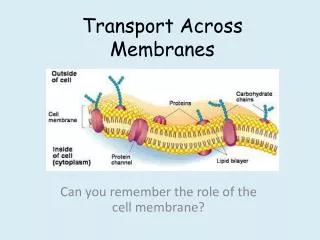
Transport across cell membranes. For the same reason that flip-flop is rare, the permeability of water, small hydrophilic molecules and ions is low. So, specific transport mechanisms are needed. Also, the differential distribution of ions and other molecules (e.g. glucose)

E N D
Transport across cell membranes For the same reason that flip-flop is rare, the permeability of water, small hydrophilic molecules and ions is low. So, specific transport mechanisms are needed. Also, the differential distribution of ions and other molecules (e.g. glucose) serves as a driving force (stored energy) So, pumps are needed to maintain these gradients.
But obviously large and polar molecules are found in cells. How do they get in??
Researchers need experimental systems in which • a particular membrane-transport protein predominates. • Heterologous expression • Liposomes
GLUT1 increases gluc. transport and is specific for D glucose
Facilitated transport (aka facilitated diffusion) occurs with the same change in free energy as diffusion. But it would go, due to activation energy. So, think of the channel or transporter as a catalyst. • 3 things distinguish a uniport from diffusion: • The rate of facilitated transport is higher than diffusion • Facilitated transport is specific • Facilitated transport has a Vmax
GLUT1 High to Low (RBC) 12 a helices 2% of RBC protein Specific for D glucose Pay attention to these arrow heads 12 glucose binding triggers conformational change. 23 glucose now facing the cytoplasm 3—4 glucose can be released to cytoplasm 45 glucose dissociation triggers return to original conformation How can this continue to run?? Answ: hexokinase
In summary: • The membrane is differentially permeable • Three kinds of transport • Transport can be studied in isolated systems • Uniport transport differs from diffusion in many ways • Transporters follow rules of enzymes
Typical ion concentrations in cells and blood cells (mM) blood (mM) K+ 139 4 Na+ 12 145 Cl- 4 116 Ca++ <0.0002 2 So, how is this maintained??
Intracellular ion environments and membrane potential Membrane potential is maintained by pumps But set by leak channels -70 mV = 200,000 V/cm High tension lines is 200,000/km Membrane potential drives the transport reactions on the membrane
Intracellular ion environments and membrane potential Read but don’t memorize the Nearnst explanation on p 587 Rather follow along my explanation of how resting potentials are established.
The distribution of ions on both sides of the impermeable barrier is set like the cell and blood. No ion flow possible so V=0 Now, open a Na+ selective channel Na+ move left until positive charges build on the left and negative charges on right V= -59 mV with respect to the right (out) Now, open a K+ selective channel Same except V= -59mV with respect to the cytoplasm. Intracellular ion environments and membrane potential
Question: How did the original concentration differences become established? Answer: pumps!!
P-class: heterotetramer, alpha is phosphorylated, Na+/K+ ATPase also Ca++ ATPase (SR of muscles), H+ATPase (lysozomes)
F- and V-class: 3 TM subunits, 5 extrinsic, no phosphointermediate, V types in plant vacuoles and lysozomes, F types in bacteria and mitochondria, F pumps used for ATP synthesis, V pumps for acidification
ABC superfamily: heterotetramer, 100’s of members, transport sugars, amino acids, peptides, CFTR
We’ll use the Ca++ ATPase as an example to study mechanism. F and ABC later • 2 Ca++ bind, note two E1’s • Binding induces a conformational change that permits phosphorylation. • P form causes the flip, note the change from high to low energy P bond • The flip changes the Ca++ binding site to low affinity. • Return to E1.
Na+/K+ ATPase maintains the intracellular Na+ and K+ in cells • Required: classic experiment 15.1 • Evidence that this pump is responsible for coupled K+/Na+ movement: • Oubain blocks the ATPase and Na+/K+ movement • Liposome reconstitution demonstrated Na+/K+ exchange • The mechanism is similar to the Ca++ ATPAse but not exactly the same • Coupled transport • Phosphate hydrolysis drives “K+ in” conformation Required: Media Connections, Biological Energy Conversions
[Na+]out ~400 mM [K+]out ~ 4-20 mM [Na+]in ~12-50 mM [K+]in ~400 mM Km Na+ ~ 0.6 mM Km K+ ~ 0.2 mM
AE1 protein, a Cl-/HCO3- antiporter is crucial to CO2 transport in RBC i.e an anion transporter. No net charge movement. Concentration only. Movement of CO2 from peripheral tissues (systemic capillaries) to lungs. Carbonic anhydrase in blood converts CO2 to water soluble bicarbonate/ . i.e CO2 is loaded into cells and carbonic acid is pumped out. Release of CO2 in the lungs because O2 drives carbonic anhydrase in reverse. CO2 in the lungs moves into RBC with Cl- exchange.
Next scenario: How do you move glucose through a cell?
Gut and kidney epithelial cells must pump glucose up their gradients (low to high) into the cells. Energy stored in the membrane potential drives this transport. 2 Na+ cotranspost with 1 glucose. This permits transport against a steep gradient.