Chapter 11 Structure of Nucleic Acids
560 likes | 1.05k Vues
Chapter 11 Structure of Nucleic Acids. Outline. How do scientists determine the primary structure of nucleic acids? What sorts of secondary structures can double-stranded DNA molecules adopt? Can the secondary structure of DNA be denatured and renatured?

Chapter 11 Structure of Nucleic Acids
E N D
Presentation Transcript
Outline • How do scientists determine the primary structure of nucleic acids? • What sorts of secondary structures can double-stranded DNA molecules adopt? • Can the secondary structure of DNA be denatured and renatured? • Can DNA adopt structures of higher complexity? • What is the structure of eukaryotic chromosomes? • Can nucleic acids be synthesized chemically? • What are the secondary and tertiary structures of RNA?
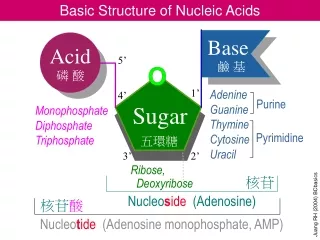
11.1 How Do Scientists Determine the Primary Structure of Nucleic Acids? • Two simple tools have made nucleic acid sequencing easier than polypeptide sequencing: • The type II restriction endonucleases that cleave DNA at specific oligonucleotide sites • Gel electrophoresis, which is capable of separating nucleic acid fragments that differ from one another in length by just a single nucleotide
11.1 How Do Scientists Determine the Primary Structure of Nucleic Acids? • Chain termination method (dideoxy method), developed by Frederick Sanger is the basis for most DNA sequencing currently. • The method takes advantage of the DNA polymerase reaction, which copies a DNA strand in complementary fashion to form a new second strand
11.1 How Do Scientists Determine the Primary Structure of Nucleic Acids? • DNA is a double-helical molecule • Each strand of the helix must be copied in complementary fashion by DNA polymerase • Each strand is a template for copying • DNA polymerase requires template and primer • Primer: an oligonucleotide that pairs with the end of the template molecule to form dsDNA • DNA polymerases add nucleotides in 5'-3' direction
11.1 How Do Scientists Determine the Primary Structure of Nucleic Acids? DNA replication yields two daughter DNA duplexes identical to the parental DNA molecule.
Chain Termination Method • Primer extension: A template DNA base-paired with a complementary primer is copied by DNA polymerase in the presence of dATP, dCTP, dGTP, dTTP • Solution contains small amounts of the four dideoxynucleotide analogs of these substrates, each of which contains a distinctive fluorescent tag, illustrated here as: • Orange for ddATP • Blue for ddCTP • Green for ddGTP • Red for ddTTP
Occasional incorporation of a dideoxynucleotide terminates further synthesis of that strand The chain termination method of DNA sequencing.
Chain Termination Method • Most of the time, the polymerase uses normal nucleotides and DNA molecules grow normally • Occasionally, the polymerase uses a dideoxynucleotide, which prevents further extension when added to the growing chain • Random insertion of dd-nucleotides leaves (optimally) at least a few chains terminated at every occurrence of a given nucleotide
Chain Termination Method • Reaction mixtures can be separated by capillary electrophoresis • Short fragments go to bottom, long fragments on top • Read the "sequence" from bottom of gel to top • Convert this "sequence" to the complementary sequence • Now read from the other end and you have the sequence you wanted - read 5' to 3'
The set of terminated strands can be separated by capillary electrophoresis
Emerging Technologies to Sequence DNA are Based on Single-Molecule Sequencing Strategies • Growing demand for sequence information is driving the development of faster and cheaper methods of DNA sequencing • Most promising are the single-molecule strategies that do not rely on Sanger-based primed synthesis of strands complementary to prepared DNA samples • One technique involves passing a single strand of DNA through a graphene monolayer pore, measuring the change in electrical conductance (ion flow) through the pore • Each base alters electrical conductance in a subtle but different way, facilitating the “reading” of sequence
Figure 11.5 DNA Sequencing through a pore in a graphene monolayer
11.2 What Sorts of Secondary Structures Can Double-Stranded DNA Molecules Adopt? The six degrees of freedom in the deoxyribose-PO4 units of the polynucleotide chain. The seventh free rotation is about the C1'-N glycosidic bond.
DNA structure (a) Double-stranded DNA as an imaginary ladderlike structure. (b) A simple right-handed twist converts the ladder to a helix.
11.2 What Sorts of Secondary Structures Can Double-Stranded DNA Molecules Adopt? • The stability of the DNA double helix is due to: • Hydrogen bonds – between base pairs • Electrostatic interactions – mutual repulsion of phosphate groups, which makes them most stable on the helix exterior • Base-pair stacking interactions • Right-twist closes the gaps between base pairs to 3.4 A (0.34 nm) in B-DNA
The “canonical” base pairs • The canonical A:T and G:C base pairs have nearly identical overall dimensions • A and T share two H bonds • G and C share three H bonds • G:C-rich regions of DNA are more stable • Polar atoms in the sugar-phosphate backbone also form H bonds
Major and minor grooves • The "tops" of the bases (as we draw them) line the "floor" of the major groove • The major groove is large enough to accommodate an alpha helix from a protein • Regulatory proteins (transcription factors) can recognize the pattern of bases and the H-bonding possibilities in the major groove
The “canonical” base pairs Watson-Crick A:T and G:C base pairs. All H-bonds in both base pairs are straight.
Double Helical Structures Can Adopt a Number of Stable Conformations • The DNA double helix can adopt several stable conformations • Helical twist is the rotation of one base pair relative to the next, around the axis of the double helix • Successive base pairs in B-DNA show a mean rotation of 36º with respect to each other • Propellor twist involves rotation around a different axis, namely an axis perpendicular to the helix axis
Double Helical Structures Can Adopt a Number of Stable Conformations (a) Helical twist: Successive base pairs in B-DNA show a rotation with respect to each other.
Double Helical Structures Can Adopt a Number of Stable Conformations Figure 11.10 Helical twist and propellor twist in DNA. (b) Propellor twist: Rotation in this dimension allows the hydrophobic surfaces of bases to overlap better
Double Helical Structures Can Adopt a Number of Stable Conformations Each of the bases in a base pair shows positive propellor twist as viewed along the N-glycosidic bond. Note how the hydrogen bonds between bases are distorted by this motion, yet remain intact.
Double Helical Structures Can Adopt a Number of Stable Conformations The B-form of the DNA double helix. In B-form, the pitch (the distance required to complete one helical turn) is 3.4 nm. Twelve base pairs of DNA are shown.
Double Helical Structures Can Adopt a Number of Stable Conformations The A-form of the DNA double helix. The pitch of the A-form helix is 2.46; thus the A-form is a shorter, wider structure than the B-form. One turn in A-form DNA requires 11 bp to complete. Twelve base pairs are shown here.
Z-DNA Discovered by Alex Rich • Found in G:C-rich regions of DNA • G goes to syn conformation • C stays anti but whole C nucleoside (base and sugar) flips 180 degrees • Result is that G:C H bonds can be preserved in the transition from B-form to Z-form!
Z-DNA is a Conformational Variation in the Form of a Left-Handed Double Helix • The Z-form of double helical DNA. • The N-glycosyl bonds of G residues in this alternating copolymer are rotated 180º with respect to their conformation in B-DNA, so now the G nucleoside is in the syn rather than the anti conformation. • The C residues remain in the anti form. • Because the G ring is flipped, the C ring must also flip to maintain normal Watson-Crick base pairing.
Comparison of A, B, Z DNA • A: right-handed, short and broad, 2.3 Å, 11 bp per turn • B: right-handed, longer, thinner, 3.32 Å, 10 bp per turn • Z: left-handed, longest, thinnest, 3.8 Å, 12 bp per turn
DNA Methylation and Epigenetics • Methylation of cytosine residues (forming 5-methylcytosine) is essential for normal embryonic development • Cytosine methylation switches genes off, so that the information they encode is not expressed • Epigenetics is the study of heritable changes in the genome that occur without a change in nucleotide sequence (such as cytosine methylation) • Epigenetic changes can influence expression of the information encoded by the genome
Intercalating Agents Distort the Double Helix • The double helix is a very dynamic structure • Because it is flexible, aromatic macrocycles – flat hydrophobic molecules composed of fused, heterocyclic rings, can slip between the stacked pairs of bases • The bases are force apart to accommodate these intercalating agents • Ethidium bromide • Acridine orange • Actinomycin D
Alternative H-Bonding Interactions Give Rise to Novel DNA Structures Cruciform structures arise from inverted repeats. In such structures, the normal interstrand base pairing is replaced by intrastrand pairing. Self-complementary inverted repeats can rearrange to form H-bonded cruciform stem-loop structures. Cruciforms are not as stable as normal DNA, because an unpaired segment must exist in the loop.
11.3 Can the Secondary Structure of DNA Be Denatured and Renatured? • When DNA is heated to 80°C or more, its UV absorbance increases by 30-40% • This hyperchromic shift reflects the unwinding of the DNA double helix • Stacked base pairs in native DNA absorb less light due to p,p electron interactions • When T is lowered, the absorbance drops, reflecting re-establishment of the double helix and base-pair stacking
The Buoyant Density of DNA Density gradient ultracentrifugation is a useful way to separate and purify nucleic acids. The net movement of solute particles in an ultracentrifuge is the result of two processes: diffusion (from regions of higher concentration to regions of lower concentration) and sedimentation due to centrifugal force.
Single-Stranded DNA Can Renature to Form DNA Duplexes • Denatured DNA will renature to re-form the duplex structure if the denaturing conditions are removed • Renaturation requires reassociation of the DNA strands into a double helix, a process termed reannealing • For this to occur, the strands must realign so that their complementary bases are once again in register and the helix can be “zippered up”
Nucleic Acid Hybridization: Different DNA Strands of Similar Sequence Can Form Hybrid Duplexes • If DNA from two different species are mixed, denatured, and allowed to cool slowly, hybrid duplexes may form, provided the DNA from one species is similar in sequence to the other • The degree of hybridization is a measure of the sequence similarity between the two species • 25% of the DNA from a human forms hybrids with mouse DNA, implying some sequence similarity • Hybridization is a common procedure in molecular biology for identifying specific genes and for revealing evolutionary relationships
11.4 Can DNA Adopt Structures of Higher Complexity? • In duplex DNA, there are ten bp per turn of helix • Circular DNA sometimes has more or less than 10 bp per turn - a supercoiled state • Enzymes called topoisomerases or gyrases can introduce or remove supercoils • Cruciforms occur in palindromic regions of DNA • Negative supercoiling may promote cruciforms
Supercoils Are One Kind of Structural Complexity in DNA Double-stranded circular DNA forms supercoils, if the strands are underwound, or overwound.
11.5 What Is the Structure of Eukaryotic Chromosomes? • Human DNA’s total length is ~2 meters! • This must be packaged into a nucleus that is about 5 micrometers in diameter • This represents a compression of more than 100,000! • It is made possible by wrapping the DNA around protein spools called nucleosomes and then packing these in helical filaments • These filaments are thought to arrange in loops associated with the nuclear matrix
Structural Organization of Chromatin Gives Rise to Chromosomes • The beads-on-a-string motif is the “primary” structure of chromatin. • The “secondary” level of chromatin structure is the 30-nm fiber, formed when an array of nucleosomes in a zig-zag pattern adopts a two-start helical conformation (Figures 11.29a, b, c). • Higher levels of chromatin structural organization are achieved when the 30-nm fiber forms long loops of 60-150,000 bp. • Electron microscopic analysis of human chromosome 4 suggests that 18 such loops are then arranged radially about the circumference of a single turn to form a miniband unit of the chromosome.
11.7 What Are the Secondary and Tertiary Structures of RNA? • The double-stranded structure of DNA imposes great constraints on its conformational possibilities • RNA molecules are typically single-stranded and thus have six to seven degrees of freedom per nucleotide unit • Thus RNA molecules have a much greater number of conformational possibilities • Complementary sequences in RNA can join via intrastrand base pairing • When the base pairing is not complete, a variety of bulges and loops can form, including hairpin stem-loop structures
11.7 What Are the Secondary and Tertiary Structures of RNA? Bulges and loops formed in RNA when aligned sequences are not fully complementary
11.7 What Are the Secondary and Tertiary Structures of RNA? • A number of defined structural motifs recur within the loops of stem-loop structures, such as U-turns, tetraloops, and bulges • Regions where several stem-loop structures meet are termed junctions • Stems, loops, bulges, and junctions are the four basic secondary structural elements in RNA • Other tertiary structural motifs arise from coaxial stacking, pseudoknot formation, and ribose zippers
11.7 What Are the Secondary and Tertiary Structures of RNA? Junctions and coaxial stacking in RNA.
11.7 What Are the Secondary and Tertiary Structures of RNA? RNA pseudoknots are formed when a single-stranded region of RNA base-pairs with a hairpin loop.
Transfer RNA Adopts Higher-Order Structure Through Intrastrand Base Pairing • In tRNA, with 73-94 nucleotides in a single chain, a majority of the bases are hydrogen- bonded to one another • Hairpin turns bring complementary stretches of bases into contact • Extensive H-bonding creates four double helical domains, three capped by loops, one by a stem • Only one tRNA structure (alone) is known • Phenylalanine tRNA is "L-shaped" • Many non-canonical base pairs found in tRNA
tRNA Tertiary Structure Arises From Interloop Base Pairing The three-dimensional structure of yeast phenylalanine tRNA. The anticodon loop is at the bottom and the acceptor end is at the top right.
Ribosomal RNA Ribosomes synthesize proteins • All ribosomes contain large and small subunits • rRNA molecules make up about 2/3 of ribosome • High intrastrand sequence complementarity leads to extensive base-pairing • Secondary structure features seem to be conserved, whereas sequence is not • There must be common designs and functions that must be conserved
Ribosomal RNA also Adopts Higher-Order Structure Through Intrastrand Base Pairing These secondary structures of several 16S rRNAs are based on computer alignment of rRNA nucleotide sequences into optimal H-bonding segments. Comparison of secondary structures of 16S-like rRNAs from several organisms.