Download

1 / 74
750 likes | 1.16k Vues
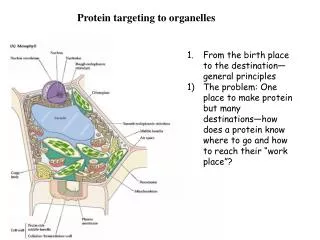
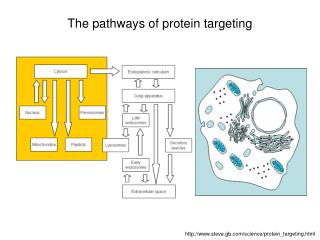
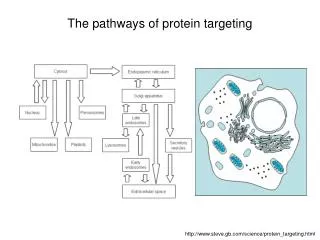
Subcellular compartments and protein targeting Alex van der Bliek. Cellular compartments. TGN. early endosome. Golgi. peroxisomes. late endosome or MVB. ERGIC. ER. lysosome. mitochondria. Compartments allow for: Specialized environments Vectorial processing

E N D
Subcellular compartments and protein targeting Alex van der Bliek
Cellular compartments TGN early endosome Golgi peroxisomes late endosome or MVB ERGIC ER lysosome mitochondria Compartments allow for: Specialized environments Vectorial processing Chemical/electrical gradients nucleus
ER Proteins are synthesized on polysomes • Free polysomes: Targeting is postranslational (mitochondria, nucleus or peroxisome) • Membrane bound: Targeting is Co-translational (ER, further sorting through Golgi) • What sends polysomes to ER membrane? • Blobel and Dobberstein (‘75) proposed the signal hypothesis • N-terminal signal (20 - 30 hydrophobic residues) targets ribosome to ER • Nascent protein is translocated into ER • Signal peptide is cleaved • How does the signal peptide send polysomes to the ER? • Walter and Blobel discovered an 11S particle: • If you strip microsomes with high KCl, then translation stops. • Add back the 11S particle and translocation resumes. • The 11S particle contains 6 different proteins and a 7S RNA. • The 11S particle was called SRP (signal recognition particle).
Experiment showing the different roles of SRP RM = rough microsomes, which have Residual SRP Preprolactin Mature prolactin No SRP, no arrest Residual SRP allows some translocation Too late for SRP K-RM = rough microsomes stripped by washing with KCl, which removes SRP Delay in adding RM causes a buildup of arrested product. Once RMs are added the arrested products are converted to mature prolactin. 70 aa leader = arrested product, consisting of signal peptide + 40 aa buried inside the ribosome Classic paper by Walter and Blobel (1981) J. Cell Biol. 91, 557-561.
SRP = 6 proteins and 1 RNA; one protein is a GTPase (SRP54) SRP receptor = heterodimer with two GTPases (SRa and SRb) SRa and SRP54 bind to each and act as others GAPs. GTP hydrolysis provides direction
Ribosome + SRP binds to SRa + b (docking proteins) Then the ribosome is passed on to the Translocase Identified by Schekman with yeast genetic screen: • Yeast with histidinol dehydrogenase in cytoplasm survive growth on plates containing histidinol. • Adding a signal sequence to histidinol dehydrogenase causes translocation of into ER. These cells can not grow on histidinol. • Mutagenize those cells and screen for mutants that now can grow on histidinol. Those mutants might have a translocation defect that keeps histidinol dehydrogenase in the cytoplasm. Result of the screen: mutants called sec61, sec62, sec63, etc. Sec61 is the translocase: a 10 pass membrane spanning protein that forms a trimer.
Proteins associated with translocation SRP Cytosol ER lumen SRa SRb sec62 sec63 sec61 TRAM BiP signal peptidase Method of protein insertion Cytoplasm C N N ER lumen N C C type I type II multipass Membrane Protein topology
Nascent polypeptide enters ER lumen and encounters: Signal peptidase cleaves signal peptide in ER lumen Nascent protein is sequestered by chaperones Hsp70 - first line of defense against aggregation, Hsp70 in the ER lumen is called BiP (binding protein)in mammals or Kar2 in yeast. BiP is essential for translocation Protein disulfide isomerase (PDI), Erp57, etc. ER lumen is oxidative -> disulfide bonds form spontaneously. Misfolded proteins need to be rearranged by PDI and ERp57 Prolyl peptide isomerase (PPI) Cis-trans isomerization of prolines is normally very slow Calnexin and calreticulin Calcium binding proteins in ER that function as chaperone., Bind and retain incompletely glycosylated proteins.
MHC Class I (antigen presentation) TAP (Transport of associated with antigen processing) Antigenic peptides made by the proteasome
to Golgi Normal processing pathway of MHC Class I ER lumen sec61 calnexin TAP mature Class I ribosome peptides proteasome viral protein Subverted pathway ER lumen US11 sec61 calnexin ribosome N-glycanase peptides ubiquitin ligase proteasome
Wahlman et al. (2007) Cell 129, 943-955. ERAD: ER associated degradation It was long thought that sec61 is involved, but in 2004 Derlin-1 and Derlin-related proteins were implicated. Is sec61 still involved? What are the minimal requirements for retrotranslocation?
Panel A: Radiolabel Fig. 1. An in vitro assay for retrotranslocation. Microsomes were preincubated with radiolabeled (35S-met) fluorescent (BOF) and glycosylation defective prepro-a-factor (pDgpaf-BOF). BOF is a fluorescent Lys-tRNA added through in vitro translation. Retrotranslocation is detected by quenching with a fluorescence quencher added to the cytosol. First, pDgpaf-BOF is imported into microsomes and the signal peptide is removed upon import (Dgpaf-BOF) Then, retrotranslocation is started by raising the temperature at 0’. Conclusions: Retrotranslocation requires cytosol. Overall decrease in radiolabel is though proteasome (Lactacystin shown in Suppl. Fig.) Panel B: Fluorescence
Fig. 2. Release of Dgpaf-BOF from microsomes with defined lumenal contents Panel A: Microsomes were extracted using high pH and resealed in the presence of Dgpaf-BOF and other defined components Panel B shows a bump, followed by gradual decrease in -cyto and -ATP, also seen with control (non-translocated) substrates, such as PDI and BiP. Panel C is corrected for this decrease. l = anti-BOF antibody, which quenches the fluorescence cpt = complete): resealed with original lumenal content. RRMs = reconstituted rough microsomes. cyto = cytosol from which haemoglobin was removed -ATP = preincubated with apyrase epoxomicin = proteasome inhibitor F = F+aBOF/F-aBOF F0 = F at 0’ Panel D: SDS gel recovers fluorescence from supernatant = direct biochemical evidence for release Conclusion: It is possible to reconstitute the reaction.Release requires cytosol and ATP.
Fig. 3. Retrotranslocation dependence on cytosolic components p97 (Cdc48), Npl4 and Ufd1 (proteins that transfer poly-ubiquitinated proteins to the proteasome) are ineffective. NEM is an alkylating agent used to inactivate the 19S particle (regulatory subunit of the proteasome) As in panel C shown on an expanded scale 26S (the complete proteasome) does work Conclusions: the proteasome is the only cytosolic component needed for retrotranslocation
Fig. 4. Retrotranslocation dependence on lumenal components Dgpaf-BOF anisotropy Panel A: some translocation occurs without lumenal content, Bip helps, but PDI is the best, just as good as complete Panel B: PDI increases Dgpaf-BOF anisotropy, suggesting that they bind, even though Dgpaf-BOF lacks cysteines Panel C: oxidized PDI works better, even though it binds more poorly to Dgpaf-BOF Wikipedia: Fluorescence anisotropy assays rotational diffusion using decorrelation of polarization in fluorescence (exciting versus emitted photons). This helps estimate the shape/size of the molecule. Conclusions: PDI is needed to fully reconstitute retrotranlocation
Fig. 5. Retrotranslocation dependence on membrane components Background: Retrotranslocation was originally thought to involve sec61, but in 2004 Derlin-1 and related proteins were shown to be involved. Does sec61 still contribute to this process? a-sec61 antibodies have no effect a-Der1 antibodies do inhibit release Expanded panel C showing that blockage occurs after a 2 min lag RNC (ribosome with nascent chain) and mutant BiP also have no effect Conclusions: Der1, but not sec61, is required for retrotranslocation.
Fig. 6. Crosslinking of Dgpaf to Derlin-1 Dgpaf was synthesized with a photoactivatable cross linker (ANB) and incoporated in RRMs. The reaction went for 0 or 15’ before UV cross linking. Samples were solubilized with detergent and coIPs were conducted with Der1, sec61 and TRAM antibodies. The image shows an autoradiogram. Arrows: 1. unidientified species 2. Dgpaf crosslinked to Der-1 3. Dgpaf alone 1 2 3 Conclusions: Dgpaf can be cross-linked with Der1, but not with sec61 or TRAM
Conclusions: On the lumenal side, PDI is enough for retrotranslocation. Derlin is required for retrotranslocation, but sec61 is not. On the cytosolic side, the proteasome is enough for retrotranslocation. Retrotranslocation complex contains derlin (reviewed in Cell 126, 237-9, 2006) but other proteins may also contribute in vivo.
Other targeting signals: Import into Mitochondria and peroxisomes ER retention
Cellular compartments TGN early endosome Golgi peroxisomes late endosome or MVB ERGIC ER lysosome mitochondria nucleus
Supernatant (cytosol) = protease sensitive Pellet (mitochondria) = protease insensitive Mitochondria (Double membrane - Related to bacteria - Endosymbiont) Mitochondria have their own DNA, transcription and translation machineries, but encode very few proteins (13 in mammals). Most mitochondrial proteins (1,500) are encoded by nuclear DNA. Synthesis is on free polysomes and import is post-translational Mitochondrial targeting signals: N-terminal amphipathic helix or internal sequences. Nascent mitochondrial leader sequences bind to MSF (mitochondrial stimulation factor), a chaperone that passes them on to the import machinery. In vitro assay for mitochondrial import (Schatz and Neupert, early 80s): + + positively charged + + a-helix N hydrophobic • - Make radiolabeled protein by in vitro transcription/translation. • Incubate with isolated mitochondria • Treat with protease • Separate by centrifugation • Analyze with SDS-PAGE
Mitochondrial protein import machinery Cytosolic Hsp70 and MSF keep the cytosolic precursors soluble. Tom70 and Tom 20 receptors for mito-leaders on the mitochondrial surface. Tom40: outer membrane import channel Tim23: inner membrane import channel Tim44, Hsp70 and GrpE drive translocation by ATP hydrolysis. Mitochondrial Hsp70, GrpE and hsp10/hsp60 facilitate folding. MPP (Mitochondrial Protein Peptidase) cleaves the mito-leader.
Wiedemann et al. 2003 Nature 424, 565-571.
Fig. 1. Mas37 deletion affects Tom40 biogenesis. Panel a: Mas37 deletion slows progression of Tom40 from precursor, through assembly complexes I and II to mature Tom complex. Assembly complex I has an aberrant size (*). Panel b: The amounts of Tom40 and porin (2 outer membrane proteins) are decreased, but the amounts of other proteins are not (IMS, inner membrane and matrix). (Western blots) BN-PAGE, autorad BN-PAGE, WB SDS-PAGE, WB Method: In vitro translate Tom40 with 35S -Met and incubate with purified mitochondria Solubilize with digitonin and analyze with Blue Native gel (BN-PAGE) and autoradiography Follow progression of Tom40 through intermediate complexes with time course.
Fig. 1. Panel c. Import of inner membrane and matrix proteins is not impaired in a mas37 deletion. Isolated mitochondria were incubated with radiolabeled precursors with or without inner membrane potential (DY)(required for import into matrix). The mitochondria were then treated with or without proteinase K to remove protein from the surface. p, precursor; I, intermediate; m, mature Fe-S, F1b, a-MPP b2D-DHFR and Su9-DHFR are all proteins imported into the matrix. AAC and PiC are inner membrane proteins. BN-PAGE shows that AAC and PiC still dimerize in a mas37 deletion. BN-PAGE, autorad SDS-PAGE, autorad
Fig. 1. Panel d. The amount of Tom complex decreases when the mas37D strains is grown at elevated temperatures. The amount of Tom40 compared with the amount of mtHsp70 (a matrix protein). (BN-PAGE, WB) (SDS-PAGE, WB) Overall conclusion from Fig. 1: Mas37D affects Tom40 import, but not or to a lesser extent import of matrix and inner membrane proteins. Imprt is halted priuot to the formation of assembly complex I.
Fig. 2. Mas37 is required for assembly of outer membrane proteins Panel a. Antibodies against Mas37 inhibit Tom40 import, but not the import of AAC or F1b AAC: BN-PAGE, autorad Tom40: BN-PAGE, autorad F1b: SDS-PAGE, autorad F1b: SDS-PAGE, quantitation
Fig. 2 Continued. Panel b. Cross-linking with EGS shows a transient interaction between Tom40 and Mas37. Mitochondria were incubated with 35S-Tom40, cross-linked at 5 or 60 min, solubilized and subjected to immuno-precipitation with mas37 antibody or preimmune serum. The red arrow points to the product of crosslinking, which exists at 5 min, but not at 60 min. Panel c. Porin assembly is also affected in mas37D mitochondria. (Porin trimerizes and then associates with several other proteins.) BN-PAGE, autorad SDS-PAGE, autorad, protease
Fig. 2. Continued. Panel d. The Mdm10 complex is also affected by Mas37D. Mdm10 is a mitochondrial outer membrane protein in a complex with Mdm12 and Mmm1. These affect mitochondrial morphology, but subsequent papers show that they are also part of the new import pathway uncovered with Mas37. Panel e. The Ugo1 complex is not affected by Mas37D. Ugo1 is in another mitochondrial outer membrane protein in a complex with Fzo1. They affect mitochondrial fusion. There does not seem to be a connection with the new import pathway uncovered with Mas37. Incubate mitochondria with35S-Mdm10 or 35S-Ugo1, solubilize the protein complex with digitonin, subject to BN-PAGE, autoradiography
Fig. 3. Mas37 is part of Assembly complex I, renamed the SAM complex Panel a. Mitochondria were incubated for 5 min with 35S-Tom40 . Assembly was allowed to continue, but halted at different stages with temperature shifts. A series of antibodies were added prior to lysis with digitonin. These antibodies cause a shift on BN-PAGE, when Tom40 is in the corresponding complex. Assembly complex I Assembly complex II Tom40 complex
Fig. 3. Continued. Panel b. SAM complex is in the mitochondrial outer membrane. Mitochondria were isolated by differential centrifugation. The outer membrane was ruptured by hypotonic swelling. Outer membrane shards reseal to form vesicles (OMV). The density of the mitochondrial outer membrane is lower than that of the inner membrane (fewer proteins). This allows for separation with a sucrose gradient. The upper panel shows WB of SDS-PAGE probed for markers for outer membrane (Tom40), intermembrane space (cyt. C), inner membrane (AAC, Tim23) and matrix (mtHsp70). The lower panel shows WB of BN-PAGE probed for the TOM and SAM complexes Conclusion: SAM and Mas37 are in outer membrane vesicles (OMV).
Fig. 3. Continued. Panel c. Effects of mutations in Tom proteins on the SAM complex. Tom22 and Tom6 deletions affect the size of the Tom complex, but Mas37 deletion does not. BN-PAGE, WB
SDS-PAGE, autorad SDS-PAGE, WB Fig. 3. Continued. BN-PAGE, WB The abnormal complex (SAM*) formed in a Tom7 mutant is protease sensitive The SAM complex is protease resistant BN-PAGE, autorad Mas37 is part of SAM and SAM* Mas37 is nevertheless cleaved by Prot K
Fig. 4. Tom40 is transported to the SAM complex via the TOM complex. Panel a: Tom mutant mitochondria are impaired in formation of assembly intermediate I, yet do not accumulate non-assembled Tom40. (Mas37D mitochondria do accumulate non-assembled Tom40) Conclusion: Tom complex is needed for the initial import. Panel b: Tom40 fragment generated by trypsin is not substantially altered in Mas37D mitochondria, suggesting that Mas37D does not affect the initial import and folding of Tom40.
Fig. 4. Continued Panel c: Accumulation of two-membrane-spanning b2D-DHFR/methotrexate, followed by import of 35S-labeled precursors and protease treatment. b2D-DHFR is the mitochondrial leader sequence of cytochrome b2 fused to DHFR. When incubated with methotrexate, the DHFR moiety becomes locked in the folded state, jamming both outer and inner membrane import channels. However, the Tom complex is 4 X more abundant than Tim complex, so porin is still imported. Hence the test with tom40-6 (a mutant allele). autorad Panel d: Protein levels of tom40-6 mitochondria. The amount of Tom complex is reduced in Tom40-6, but the steady state levels of various marker proteins are the same. WB
Fig. 4. Continued Panel e: Inhibition of Tom40 assembly by accumulated b2D-DHFR/methotrexate in energized tom40-6 mitochondria. The inner membrane potential (Dy) is need for import into the inner membrane. Blocking import with b2D-DHFR/methotrexate requires Dy. Panel f: Assembly of Tom40 in tom40-6 mitochondria is not inhibited by addition of b2D-DHFR/methotrexate in the absence of a Dy. WB BN-PAGE, autorad
General conclusions: Mas37 is part of a new import pathway for mitochondrial outer membrane proteins, such as porin and Tom40. This pathway uses the Tom complex for initial import, but then the newly imported proteins are passed on through two other complexes (Assembly complex I and II) before they are released as newly formed protein complexes. After this paper was published, another component of the assembly complexes was discovered: Sam50. Sam50 is similar to bacterial proteins that are required for insertion of b-barrel proteins in bacterial outer membranes. Sam50, porin and Tom40 are the three known b-barrel proteins in mitochondrial outer membranes. (A paper in last months JCB showed that several other mitochondrial outer membrane proteins also use the SAM pathway).
Mitochondrial import pathways for different submitochondrial destinations From a review in JCB 179, 585-591, 2007
Peroxisomes: Single membrane organelles, sequester b oxidation of fatty acids and catalase. Patients with Zellweger’s syndrome: defect in protein import (peroxisome ghosts) Mammalian cells luciferase catalase Transfected with Firefly luciferase (Subramani lab) wt Immunofluorescence catalase = control punctate = peroxisomal diffuse = cytosolic DC12 C-terminal deletions give diffuse cytosolic staining (cytoplasmic). The import signal proved to be C-terminal SKL (PTS1) A few peroxisomal proteins have anN-terminal cleaved leader(PTS2) pex mutant: Subramani identified possible receptor proteins by screening for yeast mutants that fail to grow on oleic acid. These mutants delineate import pathway.
Cellular compartments TGN early endosome Golgi peroxisomes late endosome or MVB ERGIC ER lysosome mitochondria nucleus
Back to ER Bulkflow: ER Golgi outside But some proteins stay in the ER (BiP, PDI, etc.): What keeps them there? Retrieval from cis-Golgi or exclusion from transport vesicles? ER-retention signals found by sequence comparisons: Lumenal proteins have C-terminal: -KDEL (mammals) -HDEL (S. cerevisiae) Transmembrane proteins have: -KKXX or -KXKXX Experimental evidence: transfect mammalian cells KDEL-tagged lysozyme and observe with fluorescence microscopy (wildtype lysozyme is normally secreted) wt lysozyme lysozyme + KDEL BiP DKDEL wt BiP Conclusion: KDEL is necessary and sufficient for ER retention
2nd experiment: processing of yeast prepro-a-factor • signal peptide glycosylated HDEL added • cleaved in ER cleaved in trans-Golgi • Adding HDEL to prepro-a-factor causes retention in ER. This protein is, however, • glycosylated by cis-Golgi enzymes, suggesting retrieval from the cis-Golgi. • BiP (Kar2) is secreted when HDEL prepro-a-factor is overexpressed, • suggesting the existence of an HDEL binding site that becomes saturated (= receptor??). Yeast screen for HDEL receptor Yeast cannot take up sucrose; Invertase is needed for growth on sucrose. Invertase is secreted into the periplasm and converts sucrose to fructose and glucose Add HDEL to invertase: Invertase-HDEL is stuck in ER Mutagenize the yeast and screen for colonies that grow on sucrose Analyze for ER retention defect. RESULT: ERD1 = pleitropic (no good) ERD2 = specific ER retention defect (good)
ERD2 is a 7-pass transmembrane protein Immunofluorescence shows ERD2 is primarily localized to cis-Golgi. ERD2 lies in waiting to catch ER proteins that have escaped. What sends ERD2 back to ER? Mutations in Asp197 prevent retrieval, but not binding to KDEL. This side of the helix may interact with other proteins to send ERD2 and the KDEL protein from Golgi to ER. Conclusion: ERD2 provides a mechanism for retrieval of soluble ER proteins
Transport vesicles What regulates vesicle budding? How do vesicles bud and get severed? How do vesicles find their target? How do vesicles fuse? Membrane traffic is studied with cellular, biochemical and genetic techniques. An important advance was the development of in vitro assays. These assays require: Intact organelles. Gentle lysis can be achieved by: - osmotic swelling and scraping - peeling off of membranes - slow freeze/thaw Markers, altered by transport - EndoH resistance - 3H-GlcNAc incoroporation - prepro-a-factor sedimentation
In the early 80s, James Rothman developed the “cis-medial Golgi complementation assay” Golgi isolated from glycosyltransferase mutant cell line Mutant cells express VSV-G protein, but lack the enzyme that incorporates 3H-GlcNAc Golgi isolated from wildtype cells Medial Golgi from wild type cells can complement the glycosylation defect upon successful transport 3H-GlcNAc 3H-GlcNAc Medial Golgi cis Golgi VSV-G
For successful incorporation you need • Donor (VSV-G in mutant Golgi) • Acceptor (wildtype Golgi) • Cytosol (soluble factors) • ATP • Lag-phase • Something happens • Before 3H GlcNAc • Gets incorporated • Presumably: • Vesicle budding lag-phase • Transfer and fusion • 3H-label • budding transfer fusion • medial • cis-Golgi • Classic paper by Balch et al. (1984) Cell 39, 405-416 {
acceptor incoporation Donor, ATP, yes cytosol acceptor + cytosol Donor, ATP no NEM-cytosol acceptor + cytosol Donor, ATP no Rothman’s assay was used to isolate vesicular transport proteins. NSF = NEM Sensitive Factor NEM inactivates proteins by Alkylating sulfhydrils. Mild NEM-treatment of cytosol inactivates a single protein needed for transport. NSF was the first of a series proteins involved in transport. Wildtype GTPgS GTPgS also blocks transport Small non-clathrin coated vesicles accumulate in Golgi preparations.
Method used to isolated COPs Coated vesicle formation Uncoating blocked by GTPgS Fusion blocked by NEM Bovine brain cytosol Rabbit Golgi No vesicles (donor + acceptor) Coated vesicles (+GTPgS) aCOP bCOP gCOP dCOP COPs (coat proteins) form a 600 kD complex (coatamer). The sequence of b-COP is similar to that of b-adaptin.