Download
1 / 80
800 likes | 1k Vues
CITRÁTOVÝ CYKLUS (Cyklus trikarboxylových kyselin, Krebsův cyklus). Trocha historie.

E N D
CITRÁTOVÝ CYKLUS (Cyklus trikarboxylových kyselin, Krebsův cyklus).
Trocha historie • Hans KREBS (1932) studoval rychlost oxidace malých organických molekul ledvinovou a jaterní tkání. Zjistil, že oxidovány jsou jen sukcinát, fumarát, acetát, malát a citrát. Později zjistil, že tkáně tvoří oxalacetát z pyruvátu a oxalacetát je dále oxidován. • Albert Szent-Györgyi (Maďarsko) studoval totéž na holubích létacích svalech (měřil spotřebu O2). Zjistil, že přidáním kterékoliv z výše uvedených 4 C dikarboxylových kyselin se spotřebuje mnohem více kyslíku než je třeba k oxidaci jen přidané kyseliny. Závěr: tyto sloučeniny jsou v buňce limitující. Dále zjistil, že malonát (kom. inhibitor sukcinátdehydrogenasy) inhibuje oxidaci – kritický stupeň oxidace.
Trocha historie II • Carl Martius a Franz Knoop – citrát je převáděn na isocitrát a poté na a-oxoglutarát. • Významný objev, neboť již bylo známo, že a-oxoglutarát lze enzymově převést na sukcinát. • V tomto momentě byla objasněna dráha od citrátu k oxalacetátu – ne cyklus !! • Katalytický efekt sukcinátu a dalších dikarboxylových kyselin zůstával hádankou. • H. Krebs v roce 1937 – citrát se tvoří v suspenzi svaloviny po přidání oxaloacetátu, pyruvátu nebo acetátu. • Dráha propojena v CYKLUS !! • H. Krebs se svým studentem mediciny Kurtem Henseleitem objevil již v roce 1932 – močovinový cyklus.
Nobelovy ceny • Hans Adolf KREBS, 1953 za lékařství a fyziologii spolu F. A. Lipmannem, (objev koenzymu A). • Albert von Szent-Györgyi Nagyrapolt, 1937 za lékařství a fyziologii, ("for his discoveries in connection with the biological combustion processes, with special reference to vitamin C and the catalysis of fumaric acid„).
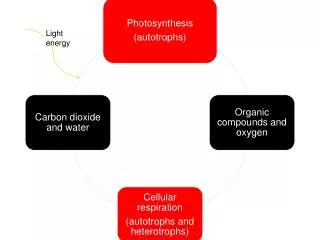
Charakteristika cyklu trikarboxylových kyselin • Lokalizován v matrix mitochondrie • Probíhá za aerobních podmínek • Amfibolický – katabolický i možnost anabolických (syntetických) pochodů. • Vstupuje dvouuhlíkatá sloučenina, která je oxidována (dehdrogenována) za tvorby redukovaných koenzymů a oxidu uhličitého. • Z cyklu se uvolňuje jediná skladovatelná energie ve formě GTP.
Tvorba acetylCoA • Z pyruvátu – produkt glykolýzy. • Z proteinů, resp. některých aminokyselin • Z acylglycerolů (tuků) – odbouráváním mastných kyselin tzv. b-oxidací. • Cyklus trikarboxylových kyselin je ústředním metabolickým dějem. • Spojují se v něm toky všech zdrojů energie a jeho komponenty slouží jako základní kameny biosyntéz různých látek jako např. sacharidů.
Tvorba acetylCoA z pyruvátu • Za aerobních podmínek je pyruvát transportován do mitochondrie výměnou za OH- pyruvátovým nosičem (aniporter). • Pyruvát je v matrix oxidativně dekarboxylován pyruvátdehydrogenasovýmkomplexem. • Pyruvát + CoA + NAD+ =acetylCoA + CO2 + NADH • Ireversibilní reakce spojuje glykolýzu a cyklus trikarboxylových kyselin.
Citrátsynthasa, EC 4. 1. 3. 7 (lyasa) • Tvorba citrátu z oxaloacetátu a acetylCoA – aldolová kondenzace • Savčí enzym je homodimer (2 x 49kd). • Poznámka: Synthasa je enzym katalyzující spojení dvou molekul (jednotek) bez spotřeby ATP (ani jiného nukleosidtrifosfátu). • Synthetasy – starý název – nyní ligasy (spojení dvou molekul za spotřeby ATP), jako např. acetylCoA-ligasa.
Prevence hydrolýzy acetylCoA • Citrátsynthasa má dva substráty, uspořádaný sekvenční mechanismus. První se váže oxalacetát, který vytvoří vazebné místo pro acetylCoA. • Katalytické místo pro hydrolýzu thioesteru se vytvoří až po tvorbě citrylCoA.
AKONITASA, EC 4. 2. 1. 3systematicky: citrát (isocitrát)hydro-lyasa • Izomerace citrátu na isocitrát přes cis-akonitát. • Akonitasa je protein obsahující železo a síru (nehemový železnatý protein). Obsahuje čtyři železnaté atomy, která jsou komplexovány čtyřmi anorganickými sulfidy (4 Fe – 4 S) a třemi cysteinovými atomy síry. Tyto komplexy prochází oxidačněredukčními procesy bez výměny protonů. Jeden atom železa váže citrát nebo isocitrát přes karboxyl nebo hydroxyl.
Isocitrátdehydrogenasa EC 1. 1. 1. 41 (první oxidačněredukční reakce cyklu). • Produkuje první sloučeninu s vysokým potenciálem přenosu - NADH.
a – Oxoglutarátdehydrogenasový komplex. • Enzymový komplex je homologní s pyruvátdehydrogenasovým. Úplně shodná je dihydrolipoyldehydrogenasová komponenta.
Sukcinyl-CoA-ligasa EC 6. 2. 1. 4 • Sukcinyl-CoA je energeticky bohatá thioesterová sloučenina. Změna standardní volné (Gibbsovy) energie je – 33, 5 kJ.mol-1. • Štěpení je spojeno s fosforylací GDP. • Savčí enzym přenáší fosfát na GDP, E. coli na GDP i ADP. • GTP je významný pro přenos signálu a dále může být nukleosiddifosfokinasou převeden na ATP. • GTP + ADP = GDP + ATP
SUKCINÁTDEHYDROGENASA EC 1. 3. 5. 1 systematicky: sukcinát:ubichinonoxidoreduktasa. • Katalyzuje oxidaci sukcinátu a ubichinonu za tvorby fumarátu a ubichinolu. • Je to železo-síra flavoprotein (FAD) • Enzym je stejně jako akonitasa protein obsahující železo a síru. Obsahuje tři druhy komplexů: 2Fe – 2S, 3Fe – 4 S, 4 Fe – 4 S. • Složen ze dvou podjednotek – heterodimer ( 70 kD a 27 kD) • Další odlišností od ostatních enzymů cyklu je, že je vázán ve vnitřní mitochondriální membráně, hlavní část komplexu II dýchacího řetězce. • Je tak přímo včleněn do elektronového transportního řetězce. • FADH2 nedisociuje z enzymu, ale je oxidován přímo kyslíkem. • Enzym je kompetitivně inhibován např. malonátem.