Download

1 / 39
390 likes | 608 Vues
Protein Dynamics Studies of Structure-Switching in Lymphotactin. By Max Shokhirev Final Project for PHAR201 Fall 2008. Overview. What is lymphotactin and why do we care? Coarse-Grained Molecular Dynamics (MD). How can we use MD to study Lymphotactin?

E N D
Protein Dynamics Studies of Structure-Switching in Lymphotactin By Max Shokhirev Final Project for PHAR201 Fall 2008
Overview • What is lymphotactin and why do we care? • Coarse-Grained Molecular Dynamics (MD). • How can we use MD to study Lymphotactin? • Creating an online user-friendly coarse-grained molecular dynamics tool to help the next generation of scientists.
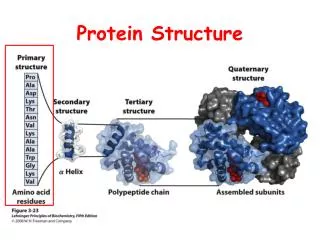
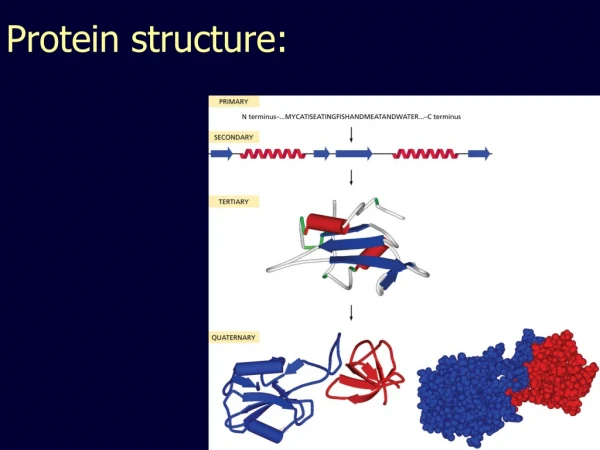
Lymphotactin is a metamorphic protein • In general an amino acid sequence determines the 3D structure of a protein • Lymphotactin has evolved to adopt two completely different tertiary folds depending on the environment it is in. • Each fold has a different function! • It is important to try to understand how and why structural switching has evolved in Lymphotactin.
Protein Folding http://upload.wikimedia.org/wikipedia/commons/thumb/a/a9/Protein_folding.png/800px-Protein_folding.png
Lymphotactin is a metamorphic protein • In general an amino acid sequence determines the 3D structure of a protein • Lymphotactin has evolved to adopt two completely different tertiary folds depending on the environment it is in. • Each fold has a different function! • It is important to try to understand how and why structural switching has evolved in Lymphotactin.
Lets take a look at the two different folds of Lymphotactin…
Overview • What is lymphotactin and why do we care? • Coarse-Grained Molecular Dynamics (MD). • How can we use MD to study Lymphotactin? • Creating an online user-friendly coarse-grained molecular dynamics tool to help the next generation of scientists.
Molecular Dynamics (MD) • Deterministic • Given initial conditions and parameters it is possible to calculate the conditions at any other point in time. • Iterative (Discrete) • Repeat force calculations at each time step and move particles accordingly. • Need to pick Δt such that the particles move continuously
Velocity-Verlet Integrator • Scheme for calculating new position, velocity, and acceleration at each time step: • Compute NewPosition • Compute Half Velocity • Compute Force • Compute Velocity Time step -1 -.5 0 .5 1 Position Velocity Acceleration
Initial Conditions… • Initial Positions • Cα positions extracted from PDB file • Even these coarse-grained models are sufficient for modeling the dynamics of small proteins (Clementi C, Nymeyer H and Onuchic J N (2000) Topological and Energetic Factors: What Determines the Structural Details of the Transition State Ensemble and ``En-route'' Intermediates for Protein Folding? An Investigation for Small Globular Proteins. J. Mol. Biol. 298: 937-953 ) • Bonding Interactions • Bonding information from PDB • Direct bonds, angles between 3 residues, dihedral angles. • Velocity? • Generated using genVel based on equipartition theory at a specified temperature. • Other parameters • Masses, LJ types, Specific LJs, general simulation parameters
Initial Temperature… • The temperature is proportional to the average speed of particles in a system. We can assign temperatures based on the Maxwell-Boltzman velocity distribution function: • Vi = (Normalized Gaussian Random number) * sqrt((Kb*Na*T)/Mi)
Temperature Control… • System is coupled to a virtual heat bath: • Vnew=Vold*sqrt(1-(ts/tau)*(1- Ttarget/Tcurrent)) • ts = time step length • tau = coupling coefficient
Force Field • Force on each particle calculated from components • Direct bond • Angle • Dihedral • Specific LJ • Non-specific LJ
Bond Interactions • V = ½k(Xi-X0)2 • Fi = k*(Xi-X0)/Xi
Lennard-Jones Interactions 10-12 LJ 6-12 LJ
Results from an example Simulation… Temperature of 350…
Results from an example Simulation… Temperature of 800…
Overview • What is lymphotactin and why do we care? • Coarse-Grained Molecular Dynamics (MD). • How can we use MD to study Lymphotactin? • Creating an online user-friendly coarse-grained molecular dynamics tool to help the next generation of scientists.
Determining Melting Temperature • Run simulation(s) at different using known structures of Ltn 10(1J8I), and Ltn 40(2JP1) • Calculate average total free energy as a function of temperature. • Specific Heat is the derivative of this function. • The peak of the specific heat curve = Tm • Need to scale the simulation to real-world values temperatures
Determining Melting Temperature • Run simulation(s) at different using known structures of Ltn 10(1J8I), and Ltn 40(2JP1) • Calculate average total free energy as a function of temperature. • Specific Heat is the derivative of this function. • The peak of the specific heat curve = Tm • Need to scale the simulation to real-world values temperatures
Alanine Scanning in silico • Which residues are important for Ltn 10 and Ltn 40 stability? • “Mutate” each of the 93 residues in each structure by removing Lennard-Jones interactions. • Measure the change in average Q value at the wildtype melting temperature
What is Q value? • Q value = Current # of Native Contacts Total # of Native Contacts
Results can be meaningful… Computed: Experimental:
Important Residues from NMR Volkman B F, Kron M A, Elgin E S, Kutlesa S, Peterson F C, Tuinistra R L (2008) Interconversion between two unrelated protein folds in the lymphotactin native state. PNAS105: 5057-5062
Is it possible to see the switching? • Unfortunately, NMR experiments performed by Volkman et al, revealed that switching was happening on the 10-1s time scale. • In order to observe the switching naturally, approximately 100 seconds of simulation time is needed. • This is not really feasible, even with coarse-grained molecular dynamics.
Steered Molecular Dynamics • We can bias the system to switching by applying an artificial force on each atom in one structure to “steer” the system toward the second form. • May or may not provide useful data.
How to simulate Dimerization? • Simulate two molecules at the same time… • Extend the forces to include forces between molecules • What keeps the molecules together? • Studies from Wolynes and coworkers have used polyglycine linkers with success as seen to the right with arc repressor Levy Y, Papoian G A, Onuchic J N, Wolynes P G (2004). Energy Landscape Analysis of Protein Dimers. Israel Journal of Chemistry. 44: 281-297
Two mutants known to stabilize one form over the other… • It has been experimentally determined that residue 55, when mutated, results in Ltn 40 being favored. On the other hand, a disulfide bond has been shown to favor Ltn 10. • Residue 55 can be “mutated” by removing or attenuating the Lennard-Jones contacts it forms • A strong disulfide bond force can be added between two Cysteine residues. • The mutants should show a shift in the system’s free energy toward one form or the other. Consequently, it is then possible to elucidate how the mutants are affecting the dynamics of Ltn.
Modeling Charges… • It has also been shown that Ltn 10 is favored in higher salt concentration environments, while Ltn 40 is dominant when salt is absent (Volkman et al). • Coulombic potential between residues. Fcoulomb = Q1Q2 r2
Significance • We live in a protein world • Proteins have evolved particular structural conformations in order to perform specific biological functions • Metamorphic proteins have evolved to adopt two or more different 3D structures depending on environmental conditions. • With the help of molecular dynamics simulation, we can take a very close look at how this structural switching is occurring in a controlled way by studying the contribution of each element in the system to the energetics of the entire system.
Overview • What is lymphotactin and why do we care? • Coarse-Grained Molecular Dynamics (MD). • How can we use MD to study Lymphotactin? • Creating an online user-friendly coarse-grained molecular dynamics tool to help the next generation of scientists.
Free Online Molecular-Dynamics Simulations… • It is needlessly difficult to study molecular dynamics. • No free user-friendly MD simulators available to the general public and scientific community • Steep learning curve • Terminal-based user interaction • Hard to set up non-standard simulations (mutations, hybrid potentials etc. ) • No integrated analysis tools
Free Online Molecular-Dynamics Simulations… • JavaMolD: a way to make MD simulations fun, interactive, and relatively easy. • Online and offline versions will be available • Simulate-Visualize-Analyze all in one application (no need to keep 12 applications open at one time!) • Define the potential by adding forces directly to the 3D structure • Built in functions to: mutate residues, merge two potentials into one (a chimera), support for Weighted Histogram Analysis of Free Energy plots. • Free to use as a research or teaching tool.
Thank you… • Feel free to email me if you have questions/comments:maxshok@gmail.com • This presentation was created as part of structural biology class taught by Dr. Philip Bourne at UCSD. Sources: Volkman B F, Kron M A, Elgin E S, Kutlesa S, Peterson F C, Tuinistra R L (2008) Interconversion between two unrelated protein folds in the lymphotactin native state. PNAS105: 5057-5062 Clementi C, Nymeyer H and Onuchic J N (2000)Topological and Energetic Factors: What Determines the Structural Details of the Transition State Ensemble and ``En-route'' Intermediates for Protein Folding? An Investigation for Small Globular Proteins. J. Mol. Biol. 298: 937-953 Shokhirev M N, Miyashita O. Molecular Dynamics Simulations using a Go-like Model to study folding of Cro protein families. (to be published in 2009) Chodera, JD, Swope, W C, Pitera, J W, Seok, C, and Dill, K A (2007).Use of the Weighted Histogram Analysis Method for the Analysis of Simulated and Parallel Tempering Simulations. J. Chem. Theory Comput.1:26 -41 Levy Y, Papoian G A, Onuchic J N, Wolynes P G (2004). Energy Landscape Analysis of Protein Dimers. Israel Journal of Chemistry. 44: 281-297