Download

1 / 37
370 likes | 387 Vues
This reference chapter provides an overview of the control mechanisms involved in transcription initiation, including alternative sigma factors, increasing initial binding of RNA polymerase, isomerization, DNA looping, proofreading, pausing, regulation through the 2˚ channel, and measurement of elongation.

E N D
Control of Transcription Initiation General References Chapter 16 of Molecular Biology of the Gene 6th Edition (2008) by Watson, JD, Baker, TA, Bell, SP, Gann, A, Levine, M, Losick, R. 547-587. Ptashne, M. and Gann, A. (2002) Genes and Signals. Cold Spring Harbor Laboratory Press, Cold Spring Harbor. Luscombe, N.M., Austin, S.E., Berman, H.M., Thornton, J.M. (2000) An overview of the structures of protein-DNA complexes. Genome Biology 1(1): reviews001.1-001.37 Examples of Control Mechanisms Alternative Sigma Factors Sorenson, MK, Ray, SS, Darst, SA (2004) Crystal structure of the flagellar sigma/anti-sigma complex 28 /FlgM reveals an intact sigma factor in an inactive conformation. Molecular Cell14:127-138. Gruber, TM, Gross, CA (2003) Multiple sigma subunits and the partitioning of bacterial transcription space. Annu. Rev. Microbiol57:441-66 Increasing the Initial Binding of RNA Polymerase Holoenzyme to DNA Lawson CL, Swigon D, Murakami KS, Darst SA, Berman HM, Ebright RH. (2004) Catabolite activator protein: DNA binding and transcription activation. Curr Opin Struct Biol. 14:10-20. Increasing the Rate of Isomerization of RNA Polymerase *Dove, S.L., Huang, F.W., and Hochschild, A. (2000) Mechanism for a transcriptional activator that works at the isomerization step. Proc Natl Acad Sci USA97: 13215-13220. Jain, D. Nickels, B.E., Sun, L., Hochschild, A., and Darst, S.A. (2004) Structure of a ternary transcription activation complex. Mol Cell 13: 45-53. Hawley and McClure (1982) Mechanism of Activation of Transcription from the l PRM promoter. JMB 157: 493-525
DNA looping **Oehler, S., Eismann, E.R., Kramer, H. and Mueller-Hill, B. (1990) The three operators of the lac operon cooperate in repression. EMBO 9:973-979. Vilar, J.M.G. and Leibler, S. (2003) DNA looping and physical constraints on transcription regulation. J Mol Biol 331:981-989. Dodd, I.B., Shearwin, K.E., Perkins, A.J., Burr, T., Hochschild, A., and Egan, J.B. (2004) Cooperativity in long-range gene regulation by the cI repressor. Genes Dev. 18:344-354. The dynamics of lac Repressor binding to its operator Elf, J., Li, G.W., and Xie, X.S. (2007). Probing transcription factor dynamics at the single-molecule level in a living cell. Science 316, 1191–1194. Li, G.W., Berg, O.G., and Elf, J. (2009). Effects of macromolecular crowding and DNA looping on gene regulation kinetics. Nat. Phys. 5, 294–297 Li, G.W., and Xie, X.S. (2011). Central dogma at the single-molecule level in living cells. Nature 475, 308–315. Hammar, P., Leroy, P., Mahmutovic, A., Marklund, E.G., Berg, O.G., and Elf, J. (2012). The lac repressor displays facilitated diffusion in living cells. Science 336, 1595–1598 *Choi, PJ, Cai,L, Frieda K and X. Sunney Xie (2008) A Stochastic Single-Molecule Event Triggers Phenotype Switching of a Bacterial Cell Science 2008: 442-446. [DOI:10.1126/science.1161427] In vivo logic of absolute rates of protein synthesis Li, GW, Burkhardt D, Gross, C and Weissman JS (2014). Quantifying absolute protein synthesis rates reveals principles underlying allocation of cellular resources. Cell.157(3):624-35. doi: 10.1016 Proofreading *Zenkin, N, Yuzenkova, y Severinov K Transcript-assisted transcriptional proofreading. Science. 2006 Jul 28;313(5786):518-20 Sydow JF, Cramer P. (2009) RNA polymerase fidelity and transcriptional proofreading.Curr Opin Struct Biol. 2009 Dec;19(6):732-9. Epub 2009 Nov 13. Sydow JF, Brueckner F, Cheung AC, Damsma GE, Dengl S, Lehmann E, Vassylyev D, Cramer P.(2009) Structural basis of transcription: mismatch-specific fidelity mechanisms and paused RNA polymerase II with frayed RNA. Mol Cell. Jun 26;34(6):710-21. Pausing Artsimovitch, I. and Landick, R (2000). Pausing by bacterial RNA polymerase is mediated by mechanistically distinct classes of signals. PNAS 97: 7090-7095
Zhang J, Palangat M, Landick R. Role of the RNA polymerase trigger loop in catalysis and pausing. Nat Struct Mol Biol. 2010 Jan;17(1):99-104. Epub 2009 Dec 6. *Shaevitz, j. Abbondanzieri E, Landick R. and Block S (2003) Backtracking by single RNA polymerase molecules observed at near base pair resolution. Nature 426: 684-687 Herbert, K., La Porta, A, Wong B, Mooney, R. Neuman, K. Landick, R. and Block, S.(2006). Sequence-Resolved Detection of Pausing by Single RNA Polymerase Molecules. Cell 125:1083-1094 *Weixlbaumer, A, Leon, K, Landick, R and Darst SA (2013) Structural basis of transcriptional pausing in bacteria. Cell. 2013 Jan 31;152(3):431-41. doi: 10.1016/j.cell.2012.12.020. Regulation through the 2˚ channel Paul BJ, Barker MM, Ross W, Schneider DA, Webb C, Foster JW, Gourse RL. (2004) DksA: a critical component of the transcription initiation machinery that potentiates the regulation of rRNA promoters by ppGpp and the initiating NTP. Cell. 6:311-22 Measurement of elongation Larson MH, Mooney RA, Peters JM, Windgassen T, Nayak D, Gross CA, Block SM, Greenleaf WJ, Landick R, Weissman JS. Science.2014: A pause sequence enriched at translation start sites drives transcription dynamics in vivo. May 30;344(6187):1042-7. Shaevitz JW, Abbondanzieri EA, Landick R, Block SM Backtracking by single RNA polymerase molecules observed at near-base-pair resolution. Nature. 2003 Dec 11;426(6967):684-7. Epub 2003 Nov 23.
Important Points • 1. Every step in transcription initiation can be regulated to increase or decrease the number of successful initiations per time. • 2. In E. coli, transcription initiation is controlled primarily by alternative factors and by a large variety of other sequence-specific DNA-binding proteins. • 3. G=RTlnKD. This means that a net increase of 1.4 kcal/mole (the approximate contribution of an additional hydrogen bond) increases binding affinity by 10-fold. Many examples of transcription activation in bacteria take advantage of such weak interactions. • 4. To activate transcription at a given promoter by increasing KB, the concentration of RNA polymerase in the cell and its affinity for the promoter must be in the range so an increase in KB makes a difference. Likewise, to activate transcription by increasing kf, the rate of isomerization must be slow enough so the increase makes a substantial difference. • 5. Network motifs give the regulatory circuit its properties • 6. Transcriptional pauses are integral to the transcription process and are extensively utilized for regulatory roles
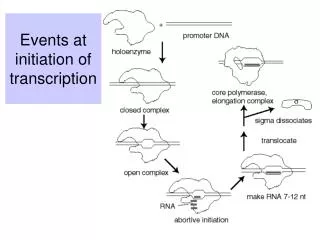
NTPs KB Kf Elongating Complex Abortive Initiation R+P RPc RPo initial binding “isomerization” Every step of transcription can be regulated DNA Binding Proteins used to alter promoter properties
Gene regulation in E. coli: The Broad Perspective • 4400 genes • 300-350 sequence-specific DNA-binding proteins • 7 factors In E. coli 1 copy/cell ≈ 10-9 M If KD = 10-9M and things are simple: 10 copies/cell 90% occupied 100 copies/cell 99% occupied
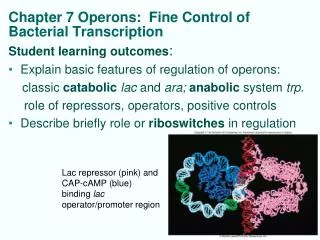
NTPs KB Kf Abortive Initiation R+P RPc RPo initial binding “isomerization” Regulation by repressors and activators Case Study: How bacteria monitor and respond to nutrient status Regulation of the lactose utilization operon: Dual negative and positive control A O P lacY lacZ lacA Repressor Activator CAP-cAMP
Lac ~ 1980 -35 -10 O3 O1 O2 Lac operator -90 -10 +400 -35 Lac 2000 Oehler, 2000 O2 1/10 affinity of O1 O3 1/300 affinity of O1 Lac repressor and DNA looping What is the function of these weak operators?
The weak operators significantly enhance represssion Oehler, 2000
Om Better! Oa M M A mutant Lac repressor that cannot form tetramers is not helped by a weak site Om Through DNA looping, Lac repressor binding to a “strong” operator (Om) can be helped by binding to a “weak” operator (OA) OK
Effects of looping (2 operators) Om (main operator) binds repressor more tightly than Oa (auxiliary operator). Transcription takes place only in the states (i) and (iii), when Om is not occupied. One operator: a single unbinding event is enough for the repressor to completely leave the neighborhood of the main operator. Two operators: repressor can escape the neighborhood of the main operator only if it sequentially unbinds both operators. Allows control of gene regulation on multiple time scales through different kinds of dissociation events Vilar, J.M.G. and Leibler, S. (2003) J Mol Biol 331:981-989 Partial dissociation: can initiate 1round of transcription (~10-20 molecules) Full dissociation: 6 min to find site again; allows establishing bistability
Positive control: activators ( e.g. CAP); facilitate RNAP binding with favorable protein-protein contact * RNAP holo A Favorable contact -35 -10 I. Activating transcription initiation at KB(initial binding) step ∆ G = RT lnKD; if * nets 1.4 kcal/mol, KB goes up 10-fold
RNAP a) If initial occupancy of promoter is low 1% occupied * A RNAP 10% occupied RNAP b) If initial occupancy of promoter is high 99% occupied * RNAP A 99.9% occupied Activating by increasing KB is effective only if initial promoter occupancy is low If favorable contact nets 1.4Kcal/mole (KB goes up 10X) then: Transcription rate increases 10-fold Little or no effect on transcription rate
cAMP high glucose How is CAP activated? Inactive CAP Active CAP Regulates >100 genes positively or negatively CAP at lac operon CAP increases transcription ~40-fold; KB ; no effect on kf A case study of activation at KB: CAP at the lac operon:
M M 1. Isolate “positive control” (pc) mutations in CAP. These mutant proteins bind DNA normally but do not activate transcription 2. “Label transfer” (in vitro) from activator labeled near putative “pc” site to RNAP S-S-X* RNAP RNAP Activate X*; reduce S-S; X* is transferred to nearest site; determine location by protein cleavage studies; X* transferred to -CTD -35 -10 M 3. Isolate CAP-non-responsive mutations in -CTD -35 -10 Strategies to identify point of contact between CAP and RNAP
Negative control: repressors (e.g. , Lac ); prevent RNAP binding R -35 -10 Positive control: activators ( e.g. CAP); facilitate RNAP binding with favorable protein-protein contact * RNAP holo A Favorable contact NTPs KB Kf Elongating Complex -35 -10 Abortive Initiation R+P RPc RPo initial binding “isomerization” Summary: Stereotypical binding of repressors and activators regulates transcription initiation
Regulatory Circuits are composed of network motifs Negative feedback loops: tunes expression to cellular state Blue line: negative feedback Red line: constant rate of A synthesis unaffected by R
Combinatorial control of gene expression AND NOT Logic, e.g. lac operon AND Logic; e.g. arabinose operon
Transcriptional pauses are really important Coordinate transcription (RNAP movement) with: 1) Folding nascent RNA 2) Other RNA processes translation, degradation, export, splicing 3) Regulator binding (TAR—HIV; RfaH prokaryotes) Promoter proximal pauses poise RNAPII for gene expression in metazoans
How to measure pauses Time (Min) Run-off transcript-- Pauses are characterized by duration and “efficiency” (probability of entering the pause state at kinetic branch between pausing and active elongation) Pause transcript-- Stall (3 NTP’s) Start reaction with 4th *NTP + heparin to prevent reinitiation Aliquots of a synchronized, radiolabeled, single-round transcription assay were removed at various times and electrophoresed on a polyacrylamide gel; separation by size Pauses can also be measured using single molecule technology
Pausing can also be measured using single molecule techniques Can follow single molecules over long times and detect very short pauses
Identification of Elemental pauses Trace of two RNA polymerase molecules, one with long pause Backtracking by eye: phase 1 (backtracking, solid line) phase 2 (pause, dotted line) phase 3 (recovery, solid line). Representative short pause (3 s); No backtracking *Short pauses account for 95% of all pausing events; subsequent studies confirmed that they are not backtracked and occur at specific sequences (ubiquitous/elemental pauses)
Pauses can also be measured genome wide using NET-seq Matt Larson ( Weissman lab)
Current view of Pausing (?) Elemental Pause Elongation Complex
Regulating Termination: Attenuation control 1. Stabilizing alternative 2˚structures of mRNA can lead to either elongation or termination 2. External inputs can alter the equilibrium between mRNA states 3. RNA polymerase pausing is critical for this regulatory mechanism
Attenuation in biosynthetic operons TAA His codons hisL hisG 1 2 3 4 No protein synthesis hisL hisG 4 3 2 1 pause hairpin transcription terminator TAA High His hisL hisG 4 3 transcription terminator 1 2 Operon mRNA level Low TAA Low His hisL hisG 3 2 High 1 4 transcription anti-terminator
Regulated “attenuation” (termination) is widespread Switch between the “antitermination” and “termination” Stem-loop structures can be mediated by: • Ribosome pausing ( reflects level of a particular charged tRNA): regulates • expression of amino acid biosynthetic operons in gram - bacteria 2. Uncharged tRNA: promotes anti-termination stem-loop in amino acyl tRNA synthetase genes in gm + bacteria 3. Proteins: stabilize either antitermination or termination stem-loop structures 4. Small molecules: aka riboswitches 5. Alternative 2˚ structures can also alter translation, self splicing, degradation
E. coli NusG: A 21kD essential elongation factor NTD CTD KOW domain NGN domain Activities: 1. Increases elongation rate 2. suppresses backtracking 3. Required for anti-termination mechanisms 4. Enhances termination mediated by the rho-factor How does one 21Kd protein mediate all of these activities?
The CTD of NusG interacts with other protein partners NusE, a ribosomal protein (S10) is part of a complex of proteins mediating antitermination/termination depending on its protein partners 50 µM NusG CTD NusE 10 nM Rho is an RNA binding hexamer that mediates termination by dissociating RNA from its complex with RNA polymerase and DNA using stepwise physical forces on the RNA derived from alternating protein conformations coupled to ATP hydrolysis Rho Although the CTD mediates the protein interactions involved in termination and antitermination, full length NusG is required for both processes, presumably because NusG must be tethered to RNA polymerase for these functions
Coupled syntheses. J W Roberts Science 2010;328:436-437 Published by AAAS
NusG, the only universal elongation factor, exhibits divergent interactions with other regulators